introduktion
den adaptive stråling af insekter på tværs af forskellige klimaer er delvis blevet lettet af den gentagne udvikling af diapause, en foregribende udviklingsarrest forud for ugunstige forhold (f.eks. vinter, tør sæson; vandhus og Norris, 1980; Tauber et al., 1986; Danks, 1987; Hodkova og Hodek, 2004). Diapause initieres typisk som reaktion på en token miljømæssig cue (f. eks., sæsonændring i fotoperiode), opretholdt i en genetisk bestemt periode og derefter afsluttet, efter at gunstige betingelser er vendt tilbage. Således tillader diapause insekter at tilpasse deres vækst og reproduktion med lokale sæsoncykler (Tauber et al., 1986; Danks, 1987). Diapause er en vigtig regulator af livshistorie, geografisk rækkevidde og sæsonbestemt overflod for mange insektbestemmere (f.eks., 2013), landbrugs skadedyr (f.eks., 1991; Li et al., 2012; Levine et al., 2015) og sygdomsvektorer (f.eks. Denlinger og Armbruster, 2014; Gray et al., 2016). Derudover påberåber diapause udbredte fysiologiske ændringer, herunder akkumulering af næringsreserver, undertrykkelse af metabolisk aktivitet, øget anaerob katabolisme, cellecyklusstop, ændringer i vnt-signalering og opregulering af forskellige stresstolerancemekanismer (Denlinger, 2002; Hahn og Denlinger, 2007, 2011; King and MacRae, 2015; Ragland and Keep, 2017). Forståelse af de reguleringsmekanismer, der ligger til grund for diapause, er et langvarigt problem i organismebiologi og vil lette udviklingen af nye ledelsesmetoder til nøglearter.
Ecdysteroider og juvenile hormoner regulerer en lang række fysiologiske og udviklingsmæssige processer i insekter (Gruntenko og Rauschenbach, 2008; Jindra et al., 2013; Palli, 2016), inklusive diapause (Denlinger, 1985; Denlinger et al., 2012). Det specifikke hormonelle signal, der regulerer diapause, har tendens til at variere afhængigt af det stadium, hvor udviklingsarrest forekommer (Denlinger et al., 2012). For eksempel er udviklingsarrest under larve eller puppe diapause typisk reguleret af ændringer i ecdysteroid overflod (f.eks., 1984; Richard et al., 1987). I modsætning hertil reguleres udviklingsarrest under voksen diapause ofte af ændringer i juvenil hormon overflod (f.eks. Spielman, 1974; Schooneveld et al., 1977; Saunders et al., 1990 og se anmeldelse i Denlinger et al., 2012).
der er et stort hul i vores forståelse af den hormonelle regulering af embryonal diapause, som er bredt observeret på tværs af Lepidoptera, Orthoptera, Hemiptera og Diptera. Tidligere forskning har i vid udstrækning dokumenteret endokrin regulering af embryonal diapause i silkeormsmølen (Bombyks mori) af diapause hormon (Hasegava, 1963; Yamashita og Hasegava, 1966; Yamashita, 1996). Imidlertid synes denne regulerende rolle for diapause hormon at være unik for B. mori og har således givet lidt indsigt i det hormonelle grundlag for embryonal diapause hos andre arter (Denlinger et al., 2012). Ud over B. mori er den hormonelle regulering af embryonal diapause blevet etableret i kun tre arter. Gypsy moth (Lymantria dispar) gennemgår en udviklingsarrest initieret og vedligeholdt af højere overflod af ecdysteroids (Lee et al., 1997; Lee og Denlinger, 1997), mens lav ecdysteroid overflod regulerer diapause i to græshopper (Chortoicetes terminifera, Locusta migratoria; Gregg et al., 1987; Jørgen et al., 2002).
til dato har ingen undersøgelse karakteriseret hormon overflod under embryonal diapause af nogen Dipteran arter. Her, vi løser dette hul ved at undersøge den hormonelle regulering af embryonal diapause i den asiatiske tiger myg, Aedes albopictus. I løbet af de sidste 30 år har denne medicinsk vigtige vektor (Paupy et al., 2009) har med succes invaderet tempererede regioner over hele verden (Benedict et al., 2007) delvis på grund af en embryonal diapause, der har lettet overlevelse under langdistancetransport (Juliano og Lounibos, 2005; Dinis et al., 2017) og udviklede sig hurtigt for at tilpasse tidspunktet for udviklingsarrest med lokale klimatiske forhold (Urbanski et al., 2012). Tempererede populationer af Ae. albopictus indtaster en maternelt reguleret, fotoperiodisk diapause. Under korte, høstlige daglængder producerer hunner æg, der fuldender embryonal udvikling, men forbliver ildfaste over for ruge stimuli som faratlarver inde i ægets korion indtil det følgende forår. I løbet af det sidste årti er transkriptionel regulering af embryonal diapause i AE. albopictus er blevet udførligt karakteriseret (Urbanski et al., 2010; Poelchau et al., 2011,2013A, b; Huang et al., 2015; et al., 2017) men den hormonelle regulering af denne afgørende tilpasning forbliver ukendt. Vi kvantificerede 20-hydroksyecdyson (20he) og juvenil hormon III (JH3) i diapausing og ikke-diapausing æg på tre tidspunkter, der beslaglægger nøglefaser af embryonal udvikling og fortolket vores resultater i sammenhæng med tidligere indsamlet Rnasek (Poelchau et al., 2013A, b) og manipulerende data (Suman et al., 2015).
materialer og metoder
ægopsamling
alle målinger anvendte F11 æg fra en laboratoriekoloni af Ae. albopictus etableret i August 2015 fra over 200 larver indsamlet i Manassas, VA, USA. Forud for dette forsøg blev befolkningen opretholdt under tidligere beskrevne betingelser . For at generere ægprøver til dette forsøg blev F10-larver opretholdt i 5,5 L Sterlitbeholdere med en densitet på cirka 250 larver pr.2.5 L deioniseret (DI) vand med 5 mL madopslæmning (Armbruster og Conn, 2006) under LD fotoperiode ved 21 liter C og 80% RH. Hver mandag–onsdag–fredag blev larver filtreret gennem et fint netnet og derefter overført til en ren 5,5 L Sterlit beholder; pupper blev fordelt til voksne bure opretholdt under enten diapause-afværgende LD fotoperiode eller diapause-inducerende, kort-dages fotoperiode (SD, 8:16 L:D).
voksne hunner fik ugentlige blodmåltider og en ovipositionsbæger foret med ubleget papirhåndklæde og halvt fyldt med DI-vand, indtil ovipositionen ophørte. Æg blev opsamlet dagligt, opretholdt på et vådt papirhåndklæde i 48 timer, derefter lufttørret og opbevaret i beholdere under SD-betingelser. 5, 7 og 11 dage efter oviposition (dpov) blev opsamlede æg vejet til nærmeste 1 liter (middelværdi: 11,460 mg, interval: 9,1–19,5 mg) på en MIKROBALANCE MH5 (Mettler-Toledo, Columbia, OH, USA), anbragt i 1,5 mL rør, snapfrosset i flydende nitrogen mellem 11 og 12 timer (tidsinterval 3-4 timer) og opbevaret ved -80 kg C. En delmængde af æg blev bevaret for at bekræfte, at ld-og SD-fotoperiodbetingelser stimulerede produktionen af henholdsvis ikke-diapause og diapause æg (se Urbanski et al., 2012).
vi samplede æg på 5, 7 og 11 dpov af to grunde. Først, Ae. albopictus indleder diapause (sensu Ko Krart Kryl, 2006; Ko krart kryl et al., 2017) efter afslutning af embryonal udvikling (Mori et al., 1981). Under de betingelser, der anvendes i vores eksperimenter, Ae. 5 dpov, og ikke-diapause-embryoner er kompetente til at klække ved cirka 7 dpov (Poelchau et al., 2013a). Ved 11 dpov er udviklingsarrest fast etableret i diapause-embryoner (Poelchau et al., 2013b). Dermed, vi valgte at sammenligne diapause og ikke-diapause embryoner på tidspunkter, der brackede perioden med udklækningskompetence, herunder etablering af udviklingsarrest under diapause. For det andet valgte vi tidspunkter, der lignede dem, der blev brugt til tidligere embryoneksperimenter (Poelchau et al., 2013A, b), så disse data kunne udnyttes til at fortolke vores nuværende resultater. Disse resultater viste sig tidligere at være godt korrelerede med genekspressionsresultater opnået via PCR (Poelchau et al., 2013a).
Ecdysteroid kvantificering
for at ekstrahere ecdysteroider blev ægprøver anbragt i et borosilikatglas 0,5 mL konisk bund mikrovævssliberør (hvede, Millville, NJ, USA) med 1 liter farnesol (Sigma-Aldrich, St. Louis, MO, USA) tilføjet som en intern standard. Æg blev manuelt homogeniseret tre gange med en Teflonpestle i 300 liter HPLC-grade kølet MeOH derefter spundet ved 12.000 RCF i 10 minutter i en nedkølet (4 liter C) centrifuge. Den resulterende supernatant blev samlet og derefter elueret gennem aluminium oksidkolonner to gange med kølet 90% MeOH og en gang med MeOH. Prøver blev tørret ned i en nedkølet (4 liter C) vakuumcentrifuge, resuspenderet i 20 liter MeOH og opbevaret ved -80 liter C indtil analyse. Positive kontroller blev fremstillet ved at tilføje 1 liter hver af 20he (Sigma-Aldrich) og farnesol til tomme rør, der ellers blev behandlet identisk. Prøveanalyse af ecdysteroidkoncentration blev udført under anvendelse af en Mikromassekvattro mikro LC-MS/MS (vand Co., Milford, MA, Amerikas Forenede Stater). Injektionsvolumener var 5,0 liter, med separationer udført ved hjælp af frikendelse UPLC BEH C18 2,1 mm med 50 mm søjler med en 1,7 liter stationær fase (farvande Co.). Driftsbetingelserne for LC-kørsler var en mobil fasestrømningshastighed på 0,37 mL min–1 med en binær mobil fase på 0,1% myresyre i acetonitril og 0,1% myresyre i vand. Indledende betingelser var 1:99 acetonitril: vand efterfulgt af isokratisk strømning i 0,3 min. Ved 0,3 min, en lineær gradient fra 1:99 Til 99:1 acetonitril:vand blev påført over 4,2 min efterfulgt af 1,0 min isokratisk strømning ved 99:1 acetonitril:vand, hvorefter den mobile fase vendte tilbage til 1:99 acetonitril:vand. Massespektrometerindstillinger var elektrospraypositive med en ødelæggelsestemperatur på 350 liter C. 20he og farnesol elueret ved henholdsvis 2,89 og 4,97 min og blev kvantificeret ved hjælp af multiple reaktionsovervågning af karakteristiske overgange: henholdsvis 481,22 (m/å) > 445,24 (m/å) og 205,41 (m/å) > 121,09 (m/å). Detektionsgrænsen for analysen er ca. 5 pg.
juvenil hormon kvantificering
hver ægprøve blev anbragt i et borosilikatglas 0,5 mL konisk bund mikrovævssliberør (hvede) med 1 liter farnesol (Sigma-Aldrich) tilføjet som en intern standard. Æg blev manuelt homogeniseret tre gange med en Teflonpestle i 500 liter HPLC-kølet sekskant, derefter spundet ved 12.000 RCF i 10 minutter i en nedkølet (4 liter C) centrifuge. Heksanfraktionerne blev rekombineret i et rent hætteglas af borosilikatglas og tørret ved vakuumcentrifugering. JH blev kvantificeret ved hjælp af gaskromatografi / massespektrometri som tidligere beskrevet (Brent og Vargo, 2003). Kort sagt blev resten vasket ud af hætteglassene med tre skylninger af geksan og tilsat til borosilikatglassøjler fyldt med aluminiumsilte. For at filtrere forurenende stoffer blev prøver elueret gennem søjlerne successivt med geksan, 10% ethylether-geksan og 30% ethylether-geksan. Efter tørring blev prøver derivatiseret ved opvarmning ved 60 liter C i 20 minutter i en opløsning af methyl-D-alkohol (Sigma-Aldrich) og trifluoreddikesyre (Sigma-Aldrich). Prøver blev tørret ned, resuspenderet i heksan, og igen elueret gennem aluminium oksidkolonner. Ikke-derivatiserede komponenter blev fjernet med 30% ethylether. JH-derivatet blev opsamlet i nye hætteglas ved tilsætning af 50% ethylacetat-geksan. Efter tørring blev prøverne resuspenderet i geksan og derefter analyseret ved hjælp af en HP 7890a-serie GC (Agilent Technologies, Santa Clara, CA, USA) udstyret med en 30 m liter 0,25 mm Bron-VOKSSØJLE (Fænomeneks, Torrance, CA, USA) koblet til en HP 5975c selektiv detektor med inert masse. Helium blev brugt som bæregas. JH-form blev bekræftet ved først at køre testprøver i scanningstilstand for kendte signaturer af JH 0, JH i, JH II, JH3, og JH3 ethyl; JH3 blev bekræftet som den primære endogene form i denne art. Efterfølgende prøver blev analyseret ved hjælp af MS SIM-tilstand, overvågning ved m / å 76 og 225 for at sikre specificitet for D3-metoksyhydrin-derivatet af JH3. Den samlede overflod blev kvantificeret mod en standardkurve for derivatiseret JH3 og justeret for ægens startmasse. Detektionsgrænsen for analysen er ca.1 pg.
statistiske analyser
analyser blev udført i R (R Core Team, 2019). JH3 overflod blev log-transformeret og analyseret ved hjælp af ANOVA til at teste for effekter af diapause status, alder, og deres interaktion. 20HAN overflod var ikke-lineær og passer ved hjælp af to polynomiske regressionsmodeller: en fuld model inklusive alder og udviklingsstatus (diapause, ikke-diapause) og en reduceret model, der kun indeholder alder. Modeller blev sammenlignet ved hjælp af ANOVA for at afgøre, om inkludering af udviklingsstatus signifikant forbedrede forklaringskraften.
Transkriptionsanalyser anvendt tidligere indsamlede data fra diapause og ikke-diapause Ae. albopictus æg (supplerende datasæt 1; Poelchau et al., 2013A, b). Differentielt udtrykte gener ved 3, 6 og 11 dpov blev undersøgt ved hjælp af gensæt berigelsesanalyser (Mootha et al., 2003) ved hjælp af Klaverpakken (V Kurrremo et al., 2013) indstillet til standardparametre efterfulgt af FDR-korrektion. Et referencegensæt blev kommenteret med de biologiske process go-termer tildelt på Vektorbase (Giraldo-Calder Larsn et al., 2015). JH3-og ecdysteroidsyntesevejsgener såvel som JH3-receptorer og nedbrydningsgener blev manuelt kommenteret i henhold til tidligere litteratur (Bai et al., 2007; Li et al., 2011; Nousova et al., 2011; Nibe og Nibe, 2014; Nibe et al., 2016). Derudover kommenterede Vi JH3-og ecdysteroid-inducerbare gener baseret på genbeskrivelser leveret i Vektorbase (Giraldo-Calder Kurstn et al., 2015). En komplet oversigt over disse gensæt findes i supplerende datasæt 1.
resultater
gennemsnitlig forekomst af diapause var 95,9% (interval: 90,5–100%) under SD-fotoperiode og 5.4% (interval: 1,8–9,5%) under LD fotoperiode (supplerende datasæt 2); således inducerede vores fotoperiodiske behandlinger de forventede diapause-reaktioner. 20HAN toppede under både diapause og ikke-diapause betingelser ved 7 dpov,men inklusive diapause status som en faktor i en kvadratisk model for 20han overflod forbedrede ikke modellens forklarende effekt (figur 1a og supplerende datasæt 3; F3, 43 = 0,21, p = 0,890). Disse resultater indikerer, at diapause-status ikke signifikant forklarer variation i 20he titer. Analysen viste, at hverken 20he-syntesevejen eller de 20he-inducerbare gener blev signifikant ændret under diapause vs. ikke-diapause betingelser ved 3, 6 eller 11 dpov (tabel 1).

tabel 1. Gensæt berigelsesanalyseresultater, se supplerende datasæt 1 for en komplet oversigt over disse gensæt.
JH3 blev bekræftet at være den primære form for juvenil hormon i denne art. JH3-overflod blev signifikant reduceret i diapause-embryoner (figur 1b og supplerende datasæt 3; F1,64 = 5,08, p = 0,028). Alder (F2,64 = 0,66, p = 0,518) og interaktionen mellem diapause-status og alder (F2,64 = 0,13, p = 0,883) påvirkede imidlertid ikke JH3-niveauerne. JH3-syntesevejen blev signifikant undertrykt ved 3 dpov, og JH-inducerbare gener blev signifikant undertrykt ved 6 dpov (figur 2 og tabel 1).
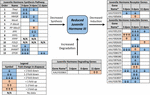
figur 2. Resume af differentiel ekspression af gener involveret i JH3-syntese, nedbrydning og induktion for Ae. albopictus (Poelchau et al., 2013A, b). Signifikante diapause-inducerede reduktioner (blå) eller stigninger (orange) er angivet med skraverede celler. Differentialstørrelsen er angivet med pile som beskrevet i legenden nederst til venstre. – Gener uden almindelige navne identificeres ved hjælp af VEKTORBASE-ID-nummer.
Diskussion
Diapause er en udbredt tilpasning, der gør det muligt for insekter at tilpasse deres vækst og reproduktion med sæsonmæssige gunstige forhold (Tauber et al., 1986; Danks, 1987). At forstå de hormonelle ændringer, hvormed insekter koordinerer opfattelsen af eksterne, diapause-inducerende signaler med de fysiologiske mekanismer, der fører til udviklingsarrest, er et langvarigt mål i biologi (Lees, 1956; Denlinger, 2002). Årtiers forskning har ført til identifikation af fælles hormonelle strategier for diapause i larve -, puppe-og voksenstadierne; den hormonelle regulering af embryonal diapause forbliver dog stort set ukendt, især i Diptera (Denlinger, 1985, 2002; Denlinger et al., 2012). I denne undersøgelse bekræftes vores direkte måling af reduceret JH3-overflod i diapause af tidligere genekspressionsprofilering (Poelchau et al., 2013A, b) og manipulerende eksperimenter (Suman et al., 2015) understøtter hypotesen om, at JH3 regulerer embryonal diapause hos denne art.
Ecdysteroids
i den foreliggende undersøgelse målte vi direkte ecdysteroid overflod på tre tidspunkter under diapause og ikke-diapause betingelser i embryoner af myggen ae. albopictus. I overensstemmelse med tidligere resultater fra Toksorhynchites amboinensis (Russo og Vestbrook, 1986) toppede 20HAN-niveauerne nær afslutningen af embryonal udvikling (7 dpov; figur 1a). Imidlertid, 20he titer adskiller sig ikke signifikant mellem diapause og ikke-diapause æg. Desuden gennemgik hverken 20he-syntesevejen eller 20he-inducerbare gener nogen signifikante, koordinerede ændringer i genekspression (tabel 1). Vi konkluderer, at I modsætning til tidligere undersøgte embryonale diapauses i Lepidoptera og Orthoptera (Hasegaja, 1963; Yamashita og Hasegaja, 1966; Gregg et al., 1987; Yamashita, 1996; Lee og Denlinger, 1997; Lee et al., 1997; Jørgen et al., 2002), 20HAN regulerer ikke diapause-initiering i Ae. albopictus.
Juvenilhormon
i modsætning hertil observerede vi ca.to gange lavere JH3-overflod i diapause-embryoner (figur 1b). Mens mindre i størrelse end forskelle i JH3 overflod påvist under diapause vs ikke-diapause betingelser i insekter, der diapause på andre livsfaser (f.eks. Yin og Chippendale, 1979; rollator og Denlinger, 1980; Readio et al., 1999), bekræftes vores direkte målinger af transkriptionelle beviser (figur 2). For eksempel undertrykkes JH3-syntesevejen signifikant under tidlig diapause-forberedelse (3 dpov). Til støtte for vores fortolkning af disse resultater har tidligere undersøgelser fundet, at reduceret ekspression af gener langs JH3-syntesevejen er stærkt korreleret med lavere JH3-overflod i begge Aedes aegypti., 2011; Rivera-Peres et al., 2014) og Diploptera punctata (Huang J. et al., 2015). JH-inducerbare gener undertrykkes også signifikant under sen diapause-forberedelse (6 dpov; tabel 1), hvilket antyder, at diapause-bestemte embryoner har en begrænset kapacitet til at reagere på JH3-signalering nær det tidspunkt, hvor ikke-diapause-embryoner bliver kompetente til at klække. Desuden genekspression for en JH3-nedværdigende (JH esterase) er signifikant opreguleret i tidlig diapause vedligeholdelse (11 dpov; figur 2) såvel som senere i diapause vedligeholdelse (21 og 40 Dpov; Poelchau et al., 2013b), der antyder, at JH3 kan forblive ved lav overflod i hele diapause hos denne art.
reduceret JH3-overflod under udviklingsarrest er noget kontraintuitivt, fordi lav juvenil hormontiter typisk er forbundet med progressive Molter, især fra larve til pupalstadium (Palli, 2016). Anvendelse af en JH3-analog (pyriproksyfen) til diapause Ae. albopictus æg accelererer hastigheden af diapause ophør på en dosisafhængig måde (Suman et al., 2015). 80% af de eksponerede æg diapause med 30 dpov sammenlignet med kun 1% diapause ophør med 80 dpov for kontrolæg (Suman et al., 2015). Ændringer i JH3-overflod kan også generere en embryonal diapause-lignende fænotype hos andre arter. For eksempel i B. mori, eksperimentel reduktion af JH3-syntese ved JHAMT knockout resulterer i fuldt udviklede faratlarver, der ikke reagerer på ruge stimuli; denne fænotype kan reddes på en dosisafhængig måde ved anvendelse af enten ekstraheret JH3 eller methopren, en JH3-analog (Nakao et al., 2015). Tilsvarende eksperimentel tilskud af en anden juvenilhormonanalog (RO-20-3600) til nyligt deponerede Drosophila melanogaster æg producerer faratlarver, der er mobile i ægget, men som ikke klækkes (Smith og Arking, 1975). Tilsammen indikerer disse data, at juvenil hormon overflod sandsynligvis bidrager til reguleringen af ruge reaktioner i forskellige insekter. Embryonal diapause i farat larvestadiet defineres af en manglende reaktion på ruge stimuli således, ændringer i juvenil hormon overflod kan repræsentere en fælles hormonel strategi til regulering af denne form for diapause.
i samspil med tidligere transkriptionelle og manipulerende data implicerer vores resultater stærkt reduceret JH3-overflod som den sandsynlige regulator for embryonal diapause i AE. albopictus. Denne forskning er den første direkte kvantificering af hormon overflod under embryonal diapause i enhver Dipteran art og repræsenterer et vigtigt skridt i retning af at afklare den hormonelle regulering af denne afgørende tilpasning til ugunstige miljøer. Karakterisering af forbindelsen mellem eksterne stimuli og udviklingsarrest via hormonel signalering er fortsat en stor udfordring (Denlinger, 1985; Denlinger et al., 2012). At belyse disse veje i en række arter vil give kritisk indsigt i grundlæggende og anvendte problemer, herunder forståelse af det molekylære og fysiologiske grundlag for livshistoriens udvikling, forudsigelse af artsreaktioner på klimaændringer og identifikation af nye mål for håndtering af skadedyrs-og vektorarter (Denlinger, 2008).
data tilgængelighed Erklæring
alle datasæt genereret til denne undersøgelse er inkluderet i artiklen/supplerende materiale.
Forfatterbidrag
SB, CB og PA bidrog til udformningen, udformningen af undersøgelsen og udarbejdede det oprindelige udkast til manuskriptet. JS, MM og JS udførte dyrehold og indsamlede ægprøverne. CB udførte LC-MS/MS og GC-MS analyserne. SB har foretaget de statistiske analyser. Alle forfattere bidrog til manuskriptrevisionen og godkendte den indsendte version.
finansiering
dette arbejde blev støttet af National Institutes of Health (1r15a|111328-01) og Davis-familiens begavelse til PA.
interessekonflikt
forfatterne erklærer, at forskningen blev udført i mangel af kommercielle eller økonomiske forhold, der kunne fortolkes som en potentiel interessekonflikt.
anerkendelser
vi takker Allan Knopf og Dan Langhorst for hjælp til måling af hormontitere. Vi takker også David Denlinger for nyttige kommentarer til tidligere versioner af manuskriptet. Brug af handelsnavne eller kommercielle produkter i denne artikel er udelukkende med det formål at give specifikke oplysninger og indebærer ikke anbefaling eller godkendelse fra det amerikanske landbrugsministerium. USDA er en lige muligheder udbyder og arbejdsgiver.
Supplerende Materiale
R Core Team (2019). R: et sprog og miljø for statistisk databehandling. Vienna: R Core Team.
Google Scholar
V. Larsen, L., Nielsen, J. og Nookaev, I. (2013). Berigelse af gensætanalysen af genomdækkende data ved at inkorporere retningsbestemmelse af genekspression og kombinere statistiske hypoteser og metoder. Nukleinsyrer Res. 41, 4378-4391. doi: 10.1093 / nar / gkt111
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar