- introductie
- materialen en methoden
- verzameling van eieren
- Ecdysteroïde kwantificering
- juveniele Hormoonkwantificatie
- statistische Analyses
- resultaten
- Diapause is een wijdverbreide aanpassing die insecten in staat stelt hun groei en voortplanting af te stemmen op seizoensgebonden gunstige omstandigheden (Tauber et al., 1986; Danks, 1987). Het begrijpen van de hormonale veranderingen waardoor insecten de waarneming van externe, diapause-inducerende signalen coördineren met de fysiologische mechanismen die leiden tot ontwikkelingsstilstand is een al lang bestaand doel in de biologie (Lees, 1956; Denlinger, 2002). Decennia van onderzoek hebben geleid tot de identificatie van gemeenschappelijke hormonale strategieën voor diapause in de larve, pop, en volwassen stadia; echter, de hormonale regulatie van embryonale diapause blijft grotendeels onbekend, met name in Diptera (Denlinger, 1985, 2002; Denlinger et al., 2012). In deze studie wordt onze directe meting van verminderde JH3 abundantie in diapause bevestigd door eerdere genexpressie profilering (Poelchau et al., 2013a, b) en manipulatieve experimenten (Suman et al., 2015) het ondersteunen van de hypothese dat JH3 embryonale diapauze reguleert bij deze soort. Ecdysteroïden
- juveniel hormoon
- Gegevensbeschikbaarheidsverklaring
- bijdragen van auteurs
- financiering
- belangenconflict
- Dankbetuigingen
- Aanvullend Materiaal
introductie
de adaptieve straling van insecten in verschillende klimaten is gedeeltelijk vergemakkelijkt door de herhaalde evolutie van de diapauze, een anticiperende ontwikkelingstilstand voorafgaand aan ongunstige omstandigheden (bijv. winter, droog seizoen; Waterhouse and Norris, 1980; Tauber et al., 1986; Danks, 1987; Hodkova and Hodek, 2004). Diapause wordt meestal gestart als reactie op een token environmental cue (bijv., seizoensgebonden verandering in fotoperiode), gehandhaafd gedurende een genetisch bepaalde periode, en vervolgens beëindigd na gunstige omstandigheden zijn teruggekeerd. Zo, diapause maakt het mogelijk insecten om hun groei en voortplanting af te stemmen op de lokale seizoensgebonden cycli (Tauber et al., 1986; Danks, 1987). Diapause is een belangrijke regulator van de levensgeschiedenis, geografisch bereik, en seizoensgebonden overvloed voor veel insect bestuivers (bijvoorbeeld, Kemp and Bosch, 2005; Fründ et al., 2013), landbouwplagen (e.g., Pullin et al., 1991; Li et al., 2012; Levine et al., 2015), en vectoren van de ziekte (e.g., Denlinger en Armbruster, 2014; Gray et al., 2016). Bovendien, de diapauze roept wijdverspreide fysiologische wijzigingen met inbegrip van accumulatie van nutriëntenreserves, afschaffing van metabolische activiteit, verhoogd anaërobe katabolisme, celcyclus arrestatie, veranderingen in WNT signaleren, en upregulation van diverse mechanismen van de spanningstolerantie aan (Denlinger, 2002; Hahn en Denlinger, 2007, 2011; King en MacRae, 2015; Ragland en houden, 2017). Inzicht in de regulerende mechanismen die ten grondslag liggen aan de diapause is een al lang bestaand probleem in de biologie van het organisme en zal de ontwikkeling van nieuwe managementbenaderingen voor belangrijke soorten vergemakkelijken.
Ecdysteroïden en juveniele hormonen regelen een breed scala aan fysiologische en ontwikkelingsprocessen bij insecten (Gruntenko and Rauschenbach, 2008; Jindra et al., 2013; Palli, 2016), met inbegrip van diapause (Denlinger, 1985; Denlinger et al., 2012). De specifieke hormonale signaal regulerende diapause neigt te variëren door het stadium waarin ontwikkelingsstilstand optreedt (Denlinger et al., 2012). Bijvoorbeeld, ontwikkelingsstilstand tijdens larve of pupale diapause wordt typisch geregeld door veranderingen in ecdysteroid abundantie (bijv., Ohtaki and Takahashi, 1972; Bowen et al., 1984; Richard et al., 1987). Daarentegen wordt de ontwikkelingsstilstand tijdens de volwassen diapauze vaak gereguleerd door veranderingen in de abundantie van juveniele hormonen (bijv. Spielman, 1974; Schooneveld et al., 1977; Saunders et al., 1990 en zie recensie in Denlinger et al., 2012).
er bestaat een groot hiaat in ons begrip van de hormonale regulatie van de embryonale diapauze, die op grote schaal wordt waargenomen bij Lepidoptera, Orthoptera, Hemiptera en Diptera. Eerder onderzoek heeft uitgebreid de endocriene regulatie van embryonale diapause in de zijderupsmot (Bombyx mori) door diapause hormoon (Hasegawa, 1963; Yamashita en Hasegawa, 1966; Yamashita, 1996). Deze regulerende rol voor diapausehormoon blijkt echter uniek te zijn voor B. mori en heeft dus weinig inzicht verschaft in de hormonale basis van embryonale diapause bij andere soorten (Denlinger et al., 2012). Voorbij B. mori, is de hormonale regulatie van embryonale diapause vastgesteld in slechts drie soorten. De gypsy moth (Lymantria dispar) ondergaat een ontwikkelingsstilstand geïnitieerd en onderhouden door een hogere overvloed van ecdysteroïden (Lee et al., 1997; Lee and Denlinger, 1997) terwijl de lage ecdysteroïde abundantie de diapauze reguleert bij twee sprinkhanen (Chortoicetes terminifera, Locusta migratoria; Gregg et al., 1987; Tawfik et al., 2002).
tot op heden heeft geen enkel onderzoek de aanwezigheid van hormonen tijdens de embryonale diapauze van enige dipteran-species gekarakteriseerd. Hier pakken we deze kloof aan door de hormonale regulatie van de embryonale diapauze te onderzoeken bij de Aziatische tijgermug, Aedes albopictus. In de afgelopen 30 JAAR, Deze medisch belangrijke vector (Paupy et al., 2009) is met succes gematigde regio ‘ s over de hele wereld binnengevallen (Benedict et al., 2007) deels te wijten aan een embryonale diapauze die overleving tijdens lange afstand transport heeft vergemakkelijkt (Juliano and Lounibos, 2005; Diniz et al., 2017) en snel geëvolueerd om de timing van de ontwikkeling arrestatie af te stemmen op de lokale klimatologische omstandigheden (Urbanski et al., 2012). Gematigde populaties van Ae. albopictus treedt in een maternaal geregelde, fotoperiodische diapauze. Bij korte, herfstachtige daglengtes produceren wijfjes eieren die de embryonale ontwikkeling voltooien, maar die tot het volgende voorjaar ongevoelig blijven voor broedprikkels als keellarven in de korion van het ei. In het afgelopen decennium, de transcriptionele regulatie van embryonale diapause in Ae. albopictus is uitgebreid gekarakteriseerd (Urbanski et al., 2010; Poelchau et al., 2011, 2013a, B; Huang X. et al., 2015; Batz et al., 2017) maar de hormonale regulatie van deze cruciale aanpassing blijft onbekend. We gekwantificeerd 20-hydroxyecdysone (20HE) en juveniele hormoon III (JH3) in diapausing en niet-diapausing eieren op drie tijdstippen dat bracket belangrijke stadia van embryonale ontwikkeling en interpreteerde onze resultaten in de context van eerder verzamelde RNAseq (Poelchau et al., 2013a, b) en manipulatieve gegevens (Suman et al., 2015).
materialen en methoden
verzameling van eieren
bij alle metingen werd gebruikgemaakt van F11 eieren uit een laboratoriumkolonie van Ae. albopictus is in augustus 2015 ontstaan uit meer dan 200 larven verzameld in Manassas, VA, Verenigde Staten. Voorafgaand aan dit experiment werd de populatie in stand gehouden onder eerder beschreven omstandigheden . Om eimonsters voor dit experiment te genereren, werden F10 larven in 5,5 l Sterliet containers gehouden met een dichtheid van ongeveer 250 larven per 2.5 L gedeïoniseerd (DI) water met 5 mL voedselslurry (Armbruster en Conn, 2006) onder LD-fotoperiode bij 21°C en 80% RV. Elke maandag–woensdag–vrijdag werden de larven door een fijnmazig net gefilterd en vervolgens overgebracht naar een schone 5.5 L sterliet container; poppen werden verdeeld in volwassen kooien onder ofwel diapause-afwendende LD fotoperiode ofwel diapause-inducerende, korte dag fotoperiode (SD, 8:16 L:D).
volwassen vrouwen kregen wekelijks bloedmaaltijd en een ovipositiebeker bekleed met ongebleekt papieren handdoek en half gevuld met DI water totdat het ovipositiebeker ophield. Eieren werden dagelijks verzameld, bewaard op een natte papieren handdoek gedurende 48 uur, vervolgens aan de lucht gedroogd en opgeslagen in containers onder SD-omstandigheden. Op 5, 7 en 11 dagen na de ovipositie (dpov) werden de verzamelde eieren gewogen tot op 1 µg (gemiddeld: 11,460 mg, bereik: 9,1-19,5 mg) op een mx5-microbalans (Mettler-Toledo, Columbia, OH, Verenigde Staten), geplaatst in buisjes van 1,5 mL, tussen 11 uur en 12 uur (Zeitgebertijd 3-4 uur) in vloeibare stikstof, en bewaard bij -80°C. Een subgroep van eieren werd behouden om te bevestigen dat LD en SD fotoperiode voorwaarden stimuleerde de productie van niet-diapause en diapause eieren, respectievelijk (zie Urbanski et al., 2012).
we hebben eieren bemonsterd bij 5, 7 en 11 dpov om twee redenen. Eerst, Ae. albopictus initieert diapause (sensu Koštál, 2006; Koštál et al., 2017) na voltooiing van embryonale ontwikkeling (Mori et al., 1981). Onder de omstandigheden gebruikt in onze experimenten, Ae. albopictus embryo ’s ondergaan segmentatie bij ongeveer 5 dpov en niet-diapause embryo’ s zijn bekwaam om uit te komen bij ongeveer 7 dpov (Poelchau et al., 2013a). Door 11 dpov, ontwikkelingsstilstand is stevig gevestigd in diapause embryo ‘ s (Poelchau et al., 2013b). Daarom hebben we ervoor gekozen om diapause en niet-diapause embryo ‘ s te vergelijken op tijdstippen tussen de periode van het uitkomen van de bekwaamheid, met inbegrip van de vestiging van ontwikkelingsstilstand tijdens diapause. Ten tweede kozen we tijdpunten die vergelijkbaar waren met die gebruikt voor eerdere RNAseq experimenten van embryo ‘ s (Poelchau et al., 2013a, b) zodat deze gegevens kunnen worden gebruikt om onze huidige resultaten te interpreteren. Deze rnaseq resultaten werden eerder gevonden goed gecorreleerd met genexpressie resultaten verkregen via qRT-PCR (Poelchau et al., 2013a).
Ecdysteroïde kwantificering
om ecdysteroïden te extraheren, werden eimonsters geplaatst in een borosilicaatglas 0,5 mL conische bodem micro weefselslijpbuis (Wheaton, Millville, NJ, Verenigde Staten) met toevoeging van 1 µg farnesol (Sigma-Aldrich, St.Louis, MO, Verenigde Staten) als interne standaard. Eieren werden drie keer manueel gehomogeniseerd met een teflon stamper in 300 µL HPLC-grade gekoelde MeOH en vervolgens gesponnen bij 12.000 RCF gedurende 10 minuten in een gekoelde (4°C) centrifuge. Het resulterende supernatant werd samengevoegd en vervolgens tweemaal geëlueerd door aluminiumoxidekolommen met 90% gekoeld MeOH en eenmaal met MeOH. De monsters werden gedroogd in een gekoelde (4°C) vacuümcentrifuge, geresuspendeerd in 20 µL MeOH, en bewaard bij -80°C tot de analyse. Positieve controles werden bereid door 1 µg elk van 20HE (Sigma-Aldrich) en farnesol toe te voegen aan lege buizen die anders identiek werden verwerkt. Monsteranalyse van de ecdysteroïdeconcentratie werd uitgevoerd met behulp van een Micromass Quattro Micro LC-MS / MS (Waters Co., Milford, Verenigde Staten). Injectievolumes waren 5,0 µL, met scheidingen uitgevoerd met ACQUITY UPLC BEH C18 2,1 mm bij 50 mm kolommen met een 1,7 µm stationaire fase (Waters Co.). De bedrijfsomstandigheden voor LC-runs waren een mobiele fasestroom van 0,37 mL min-1 met een binaire mobiele fase van 0,1% mierenzuur in acetonitril en 0,1% mierenzuur in water. De aanvankelijke voorwaarden waren 1: 99 acetonitril: water, gevolgd door isocratische stroom voor 0,3 min. Bij 0,3 min, een lineair verloop van 1:99 tot 99: 1 acetonitril: water werd gedurende 4,2 min toegepast, gevolgd door 1,0 min isocratische stroom bij 99: 1 acetonitril:water, waarna de mobiele fase terugkeerde naar 1:99 acetonitril:water. Massa spectrometer instellingen waren electrospray positief, met een desolvation temperatuur van 350°C. 20HIJ en farnesol geëlueerd op 2.89 en 4.97 min, respectievelijk, en werden gekwantificeerd aan de hand van multiple reaction monitoring van karakteristieke overgangen: 481.22 (m/z) > 445.24 (m/z), en 205.41 (m/z) > 121.09 (m/z), respectievelijk. De detectiegrens van de assay is ongeveer 5 pg.
juveniele Hormoonkwantificatie
elk eimonster werd geplaatst in een borosilicaatglas 0,5 mL microweefselmolen met conische bodem (Wheaton), waaraan als interne standaard 1 µg farnesol (Sigma-Aldrich) is toegevoegd. Eieren werden drie keer manueel gehomogeniseerd met een teflon stamper in 500 µL HPLC-kwaliteit gekoeld hexaan, vervolgens gesponnen bij 12.000 RCF gedurende 10 minuten in een gekoelde (4°C) centrifuge. De hexaanfracties werden gerecombineerd in een schone borosilicaat glazen flacon en gedroogd door vacuümcentrifugering. JH werd gekwantificeerd met behulp van gaschromatografie/massaspectrometrie zoals eerder beschreven (Brent and Vargo, 2003). Kort, het residu werd gewassen uit de flesjes met drie spoelingen hexaan en toegevoegd aan borosilicaat glazen kolommen gevuld met aluminiumoxide. Om verontreinigingen uit te filteren werden monsters achtereenvolgens geëlueerd met hexaan, 10% ethylether-hexaan en 30% ethylether-hexaan. Na droging werden de monsters afgeleid door 20 minuten bij 60°C te verhitten in een oplossing van methyl-d-alcohol (Sigma-Aldrich) en trifluorazijnzuur (Sigma-Aldrich). Monsters werden gedroogd, resuspendeerd in hexaan, en opnieuw geëlueerd door aluminium oxide kolommen. Niet-afgeleide componenten werden verwijderd met 30% ethylether. Het JH-derivaat werd verzameld in nieuwe flesjes door toevoeging van 50% ethyl-acetaat-hexaan. Na het drogen werden de monsters geresuspendeerd in hexaan en vervolgens geanalyseerd met behulp van een HP 7890A serie GC (Agilent Technologies, Santa Clara, CA, Verenigde Staten) uitgerust met een 30 m × 0,25 mm zebron ZB-WAX kolom (Phenomenex, Torrance, CA, Verenigde Staten) gekoppeld aan een HP 5975C inert mass selective detector. Helium werd gebruikt als draaggas. De JH-vorm werd bevestigd door eerst testmonsters in SCAN-modus uit te voeren voor bekende kenmerken van JH 0, JH I, JH II, JH3 en JH3 ethyl; JH3 werd bevestigd als de primaire endogene vorm bij deze soort. Daaropvolgende monsters werden geanalyseerd met behulp van de MS SIM-modus, monitoring bij M / z 76 en 225 om de specificiteit voor het D3-methoxyhydrinederivaat van JH3 te waarborgen. De totale abundantie werd gekwantificeerd aan de hand van een standaardkromme van afgeleid JH3 en gecorrigeerd voor de beginmassa van eieren. De detectiegrens van de assay is ongeveer 1 pg.
statistische Analyses
Analyses werden uitgevoerd in R (R Core Team, 2019). De overvloed van JH3 werd log-getransformeerd en geanalyseerd gebruikend ANOVA om voor gevolgen van diapausestatus, leeftijd, en hun interactie te testen. 20de abundantie was niet-lineair en fit met behulp van twee veelterm regressiemodellen: een volledig model met leeftijd en ontwikkelingsstatus (diapauze, niet-diapauze), en een verminderd model met alleen leeftijd. Modellen werden vergeleken met behulp van ANOVA om te bepalen of Inclusief ontwikkelingsstatus significant verbeterde verklarend vermogen.
transcriptionele analyses maakten gebruik van eerder verzamelde RNAseq-gegevens uit diapause en non-diapause Ae. albopictus eieren (aanvullende Dataset 1; Poelchau et al., 2013a, b). Differentieel tot expressie gebrachte genen bij 3, 6 en 11 dpov werden onderzocht door gen set verrijkingsanalyses (Mootha et al., 2003) met behulp van het Pianopakket (Väremo et al., 2013) ingesteld op standaardparameters gevolgd door FDR-correctie. Een referentie gen set werd geannoteerd met de biologische proces GO termen toegewezen op VectorBase (Giraldo-Calderón et al., 2015). JH3 en ecdysteroïde synthese pathway genen evenals JH3 receptoren en degradatie genen werden handmatig geannoteerd volgens eerdere literatuur (Bai et al., 2007; Li et al., 2011; Nouzova et al., 2011; Niwa en Niwa, 2014; Matthews et al., 2016). Bovendien annoteerden we JH3 en ecdysteroïde-induceerbare genen op basis van genbeschrijvingen die in VectorBase (Giraldo-Calderón et al., 2015). Een volledig overzicht van deze genensets wordt gegeven in aanvullende Dataset 1.
resultaten
De gemiddelde incidentie van diapause was 95,9% (bereik: 90,5–100%) onder SD-fotoperiode en 5.4% (bereik: 1,8–9,5%) onder LD fotoperiode (aanvullende Dataset 2); zo induceerden onze fotoperiodische behandelingen de verwachte diapause responses. 20de piek onder zowel diapause en niet-diapause voorwaarden bij 7 dpov maar met inbegrip van diapause status als een factor in een kwadratisch model voor 20HE abundantie niet verbetering van de verklarende kracht van het model (figuur 1A en aanvullende Dataset 3; F3,43 = 0,21, p = 0,890). Deze resultaten wijzen erop dat de diapause status de variatie in 20HE titer niet significant verklaart. Pathway analyse van RNAseq gaf aan dat noch de 20he syntheseroute noch de 20he-induceerbare genen significant veranderd waren onder diapause versus niet-diapause condities bij 3, 6 of 11 dpov (Tabel 1).

Tabel 1. Resultaten van de verrijkingsanalyse van de genensets, zie aanvullende gegevensset 1 voor een volledige samenvatting van deze genensets.
JH3 werd bevestigd om de primaire vorm van juveniele hormoon in deze species te zijn. JH3 abundantie was significant verminderd in diapause embryo ‘ s (Figuur 1B en aanvullende Dataset 3; F1,64 = 5,08, p = 0,028). Leeftijd (F2, 64 = 0,66,p = 0,518) en de interactie tussen diapause status en leeftijd (F2, 64 = 0,13,p = 0,883) hadden echter geen invloed op de JH3-spiegels. Pathway-analyse van rnaseq-gegevens gaf aan dat de JH3-syntheseroute significant werd onderdrukt bij 3 dpov en JH-induceerbare genen significant werden onderdrukt bij 6 dpov (Figuur 2 en Tabel 1).
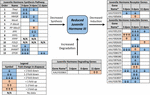
Figuur 2. Samenvatting van differentiële expressie van genen betrokken bij JH3 synthese, degradatie, en inductie voor Ae. albopictus (Poelchau et al., 2013a, b). Significante door diapauze geïnduceerde reducties (blauw) of verhogingen (oranje) worden aangegeven door gearceerde cellen. De differentiële magnitude wordt aangegeven met pijlen zoals beschreven in de legenda linksonder. *Genen zonder gemeenschappelijke namen worden geïdentificeerd door VECTORBASE ID-nummer.
Diapause is een wijdverbreide aanpassing die insecten in staat stelt hun groei en voortplanting af te stemmen op seizoensgebonden gunstige omstandigheden (Tauber et al., 1986; Danks, 1987). Het begrijpen van de hormonale veranderingen waardoor insecten de waarneming van externe, diapause-inducerende signalen coördineren met de fysiologische mechanismen die leiden tot ontwikkelingsstilstand is een al lang bestaand doel in de biologie (Lees, 1956; Denlinger, 2002). Decennia van onderzoek hebben geleid tot de identificatie van gemeenschappelijke hormonale strategieën voor diapause in de larve, pop, en volwassen stadia; echter, de hormonale regulatie van embryonale diapause blijft grotendeels onbekend, met name in Diptera (Denlinger, 1985, 2002; Denlinger et al., 2012). In deze studie wordt onze directe meting van verminderde JH3 abundantie in diapause bevestigd door eerdere genexpressie profilering (Poelchau et al., 2013a, b) en manipulatieve experimenten (Suman et al., 2015) het ondersteunen van de hypothese dat JH3 embryonale diapauze reguleert bij deze soort.
Ecdysteroïden
in deze studie hebben we direct ecdysteroïden abundantie gemeten op drie tijdstippen onder diapause en niet-diapause omstandigheden in embryo ‘ s van de mug Ae. albopictus. In overeenstemming met eerdere resultaten van Toxorhynchites amboinensis (Russo and Westbrook, 1986), piekten 20HE concentraties bijna de conclusie van embryonale ontwikkeling (7 dpov; figuur 1A). Echter, 20HE titer niet significant verschillen tussen diapause en niet-diapause eieren. Bovendien ondergingen noch de 20he-syntheseroute noch de 20he-induceerbare genen significante, gecoördineerde veranderingen in genexpressie (Tabel 1). We concluderen dat, in tegenstelling tot eerder onderzochte embryonale diapauses bij Lepidoptera en Orthoptera (Hasegawa, 1963; Yamashita en Hasegawa, 1966; Gregg et al., 1987; Yamashita, 1996; Lee and Denlinger, 1997; Lee et al., 1997; Tawfik et al., 2002), 20he regelt niet de start van de diapauze in Ae. albopictus.
juveniel hormoon
in diapauzembryo ‘ s werd daarentegen een ongeveer tweevoudige abundantie van JH3 waargenomen (figuur 1B). Hoewel kleiner in omvang dan verschillen in JH3 abundantie gedetecteerd onder diapause VS. niet-diapause voorwaarden in insecten die diapause in andere levensstadia (bijv., Yin en Chippendale, 1979; Walker en Denlinger, 1980; Readio et al., 1999), worden onze directe metingen bevestigd door transcriptioneel bewijs (Figuur 2). Bijvoorbeeld, wordt de JH3 syntheseweg beduidend onderdrukt tijdens vroege diapausevoorbereiding (3 dpov). Het ondersteunen van onze interpretatie van deze resultaten, eerdere studies hebben aangetoond dat verminderde expressie van genen langs de JH3 synthese route is sterk gecorreleerd met lagere JH3 abundantie in zowel Aedes aegypti (Nouzova et al., 2011; Rivera-Perez et al., 2014) en Diploptera punctata (Huang J. et al., 2015). JH-induceerbare genen worden ook significant onderdrukt tijdens de late diapause voorbereiding (6 dpov; Tabel 1), wat suggereert dat diapause-bestemde embryo ’s hebben een beperkte capaciteit om te reageren op JH3 signalering in de buurt van de tijd dat niet-diapause embryo’ s Bekwaam worden om uit te komen. Verder wordt genexpressie voor een JH3-degraderend enzym (JH esterase) significant verhoogd in vroege diapauseonderhoud (11 dpov; Figuur 2) evenals later in diapauseonderhoud (21 en 40 Dpov; Poelchau et al., 2013b) wat suggereert dat JH3 in de diapauze bij deze soort in geringe mate aanwezig kan blijven.
verminderde JH3-abundantie tijdens ontwikkelingsstilstand is enigszins contra-intuïtief omdat een lage juveniele hormoontiter typisch geassocieerd wordt met progressieve vervelling, met name van de larve tot het popstadium (palli, 2016). Echter, toepassing van een JH3 analoog (pyriproxyfen) diapause Ae. albopictus eieren versnelt de snelheid van de beëindiging van de diapauze op een dosisafhankelijke manier (Suman et al., 2015). Bij de meest effectieve dosering, ongeveer 80% van pyriproxyfen blootgestelde eieren beëindigen diapause met 30 dpov in vergelijking met slechts 1% diapause beëindiging met 80 dpov voor controle-eieren (Suman et al., 2015). De veranderingen in JH3 overvloed kunnen ook een embryonaal diapause-als fenotype in andere species produceren. Bijvoorbeeld in B. Mori, experimentele reductie van JH3 synthese door jhamt knockout resulteert in volledig ontwikkelde farate larven die niet reageren op broedprikkels; dit fenotype kan worden gered op een dosisafhankelijke manier door toepassing van ofwel geëxtraheerd JH3 of methopreen, een JH3 analoog (Nakao et al., 2015). Ook de experimentele aanvulling van een ander juveniel hormoon analoog (RO-20-3600) op Nieuw gedeponeerde Drosophila melanogaster eieren produceert farate larven die mobiel zijn binnen het ei, maar niet uitkomen (Smith and Arking, 1975). Samen geven deze gegevens aan dat de aanwezigheid van juveniele hormonen waarschijnlijk bijdraagt aan de regulatie van de uitbroedingsreacties bij diverse insecten. Embryonale diapauze in de farate larve stadium wordt gedefinieerd door een niet te reageren op uitbroedende stimuli dus, veranderingen in van juveniele hormoon overvloed kan een gemeenschappelijke hormonale strategie vertegenwoordigen voor het reguleren van deze vorm van diapauze.
in overeenstemming met eerdere transcriptionele en manipulatieve gegevens impliceren onze resultaten een verminderde JH3-abundantie als de waarschijnlijke regulator van embryonale diapauze bij Ae. albopictus. Dit onderzoek is de eerste directe kwantificering van hormoonovervloed tijdens embryonale diapauze in om het even welke dipteran species en vertegenwoordigt een belangrijke stap naar het verduidelijken van de hormonale regelgeving van deze cruciale aanpassing aan ongunstige milieu ‘ s. Het karakteriseren van de koppeling tussen externe stimuli en ontwikkelingsstilstand via hormonale signalering blijft een grote uitdaging (Denlinger, 1985; Denlinger et al., 2012). Het ophelderen van deze routes in een verscheidenheid van soorten zal kritisch inzicht bieden voor fundamentele en toegepaste problemen, waaronder het begrijpen van de moleculaire en fysiologische basis van de evolutie van de levensgeschiedenis, het voorspellen van species responses op klimaatverandering, en het identificeren van nieuwe doelen voor het beheer van pest-en vectorsoorten (Denlinger, 2008).
Gegevensbeschikbaarheidsverklaring
alle voor deze studie gegenereerde datasets zijn opgenomen in het artikel/aanvullend materiaal.
bijdragen van auteurs
ZB, CB en PA hebben bijgedragen aan het ontwerp en de opzet van de studie en het oorspronkelijke ontwerp van het manuscript voorbereid. ZB, MM en JS voerden de veehouderij uit en verzamelden de eimonsters. CB heeft de LC-MS/MS-en GC-MS-analyses uitgevoerd. ZB heeft de statistische analyses uitgevoerd. Alle auteurs hebben bijgedragen aan de herziening van het manuscript en hebben de ingediende versie goedgekeurd.
financiering
dit werk werd gesteund door de National Institutes of Health (1R15A|111328-01) en de familie Davis aan PA.
belangenconflict
de auteurs verklaren dat het onderzoek werd uitgevoerd zonder enige commerciële of financiële relatie die als een potentieel belangenconflict kon worden opgevat.
Dankbetuigingen
Wij danken Allan Knopf en Dan Langhorst voor hun hulp bij het meten van de hormoontiters. We danken ook David Denlinger voor nuttige opmerkingen over eerdere versies van het manuscript. Het gebruik van handelsnamen of commerciële producten in dit artikel is uitsluitend bedoeld voor het verstrekken van specifieke informatie en impliceert geen aanbeveling of goedkeuring door het Amerikaanse Ministerie van Landbouw. USDA is een aanbieder van gelijke kansen en werkgever.
Aanvullend Materiaal
R Kernteam (2019). R: een taal en omgeving voor statistische gegevensverwerking. Wenen: R Core Team.
Google Scholar
Väremo, L., Nielsen, J., en Nookaew, I. (2013). Het verrijken van de genreeks analyse van genoom-brede gegevens door directionaliteit van genuitdrukking op te nemen en statistische hypothesen en methodes te combineren. Nucleïnezuren Res. 41, 4378-4391. doi: 10.1093 / nar / gkt111
PubMed Abstract / CrossRef Full Text / Google Scholar