- Introducere
- materiale și metode
- colectarea ouălor
- ecdysteroid cuantificare
- cuantificarea hormonului Juvenil
- analize statistice
- rezultate
- discuție
- Ecdysteroids
- hormonul Juvenil
- Declarație privind disponibilitatea datelor
- contribuțiile autorului
- finanțare
- Conflict de interese
- mulțumiri
- Material Suplimentar
Introducere
radiația adaptivă a insectelor în diverse climate a fost facilitată parțial de evoluția repetată a diapauzei, o arestare anticipativă a dezvoltării înainte de condițiile nefavorabile (de exemplu, iarna, sezonul uscat; Waterhouse și Norris, 1980; Tauber și colab., 1986; Danks, 1987; Hodkova și Hodek, 2004). Diapauza este de obicei inițiată ca răspuns la un semn de mediu (de ex., schimbarea sezonieră a fotoperioadei), menținută pentru o perioadă determinată genetic și apoi încheiată după revenirea condițiilor favorabile. Astfel, diapause permite insectelor să-și alinieze creșterea și reproducerea cu ciclurile sezoniere locale (Tauber și colab., 1986; Danks, 1987). Diapause este un regulator cheie al istoriei vieții, al gamei geografice și al abundenței sezoniere pentru mulți polenizatori de insecte (de exemplu, Kemp și Bosch, 2005; fr., 2013), dăunători agricoli (de exemplu, Pullin și colab., 1991; Li și colab., 2012; Levine și colab., 2015) și vectorii bolii (de exemplu, Denlinger și Armbruster, 2014; Gray și colab., 2016). În plus, diapause invocă modificări fiziologice pe scară largă, inclusiv acumularea de rezerve de nutrienți, suprimarea activității metabolice, creșterea catabolismului anaerob, stoparea ciclului celular, modificări ale semnalizării Wnt și reglarea în sus a diferitelor mecanisme de toleranță la stres (Denlinger, 2002; Hahn și Denlinger, 2007, 2011; King și MacRae, 2015; Ragland și Keep, 2017). Înțelegerea mecanismelor de reglementare care stau la baza diapauzei este o problemă de lungă durată în biologia organismului și va facilita dezvoltarea de noi abordări de gestionare a speciilor cheie.
Ecdisteroizii și hormonii juvenili reglează o gamă largă de procese fiziologice și de dezvoltare la insecte (Gruntenko și Rauschenbach, 2008; Jindra și colab., 2013; Palli, 2016), inclusiv diapause (Denlinger, 1985; Denlinger și colab., 2012). Semnalul hormonal specific care reglează diapauza tinde să varieze în funcție de stadiul în care are loc stoparea dezvoltării (Denlinger și colab., 2012). De exemplu, arestarea dezvoltării în timpul diapauzei larvare sau pupale este de obicei reglementată de modificări ale abundenței ecdisteroide (de exemplu, Ohtaki și Takahashi, 1972; Bowen și colab., 1984; Richard și colab., 1987). În schimb, arestarea dezvoltării în timpul diapauzei adulte este adesea reglementată de modificări ale abundenței hormonilor juvenili (de exemplu, Spielman, 1974; Schooneveld și colab., 1977; Saunders și colab., 1990 și vezi recenzia în Denlinger și colab., 2012).
există un decalaj major în înțelegerea noastră a reglării hormonale a diapauzei embrionare, care este observată pe scară largă în Lepidoptera, Ortoptera, Hemiptera și Diptera. Cercetările anterioare au documentat pe larg reglementarea endocrină a diapauzei embrionare în molia viermilor de mătase (Bombyx mori) de către hormonul diapauzei (Hasegawa, 1963; Yamashita și Hasegawa, 1966; Yamashita, 1996). Cu toate acestea, acest rol de reglementare pentru hormonul diapauzei pare a fi unic pentru B. mori și, prin urmare, a oferit puține informații despre baza hormonală a diapauzei embrionare la alte specii (Denlinger și colab., 2012). Dincolo de B. mori, reglarea hormonală a diapauzei embrionare a fost stabilită la doar trei specii. Molia țigănească (Lymantria dispar) suferă o arestare de dezvoltare inițiată și menținută de o abundență mai mare de ecdisteroizi (Lee și colab., 1997; Lee și Denlinger, 1997) în timp ce abundența ecdisteroidă scăzută reglează diapauza în două lăcuste (Chortoicetes terminifera, Locusta migratoria; Gregg și colab., 1987; Tawfik și colab., 2002).
până în prezent, niciun studiu nu a caracterizat abundența hormonilor în timpul diapauzei embrionare a oricărei specii Dipterane. Aici, abordăm acest decalaj prin investigarea reglării hormonale a diapauzei embrionare la țânțarul tigru asiatic, Aedes albopictus. În ultimii 30 de ani, acest vector important din punct de vedere medical (Paupy și colab., 2009) a invadat cu succes regiunile temperate din întreaga lume (Benedict și colab., 2007) datorită parțial unei diapause embrionare care a facilitat supraviețuirea în timpul transportului pe distanțe lungi (Juliano și Lounibos, 2005; Diniz și colab., 2017) și a evoluat rapid pentru a alinia momentul opririi dezvoltării cu condițiile climatice locale (Urbanski și colab., 2012). Populațiile Temperate ale Ae. albopictus intră într-o diapauză fotoperiodică reglementată matern. În zilele scurte, de toamnă, femelele produc ouă care completează dezvoltarea embrionară, dar rămân refractare la stimulii de incubație ca larve farate în interiorul corionului oului până în primăvara următoare. În ultimul deceniu, reglementarea transcripțională a diapauzei embrionare în Ae. albopictus a fost caracterizat pe larg (Urbanski și colab., 2010; Poelchau și colab.,2011, 2013a, b; Huang X. și colab., 2015; Batz și colab., 2017), dar reglementarea hormonală a acestei adaptări cruciale rămâne necunoscută. Am cuantificat 20-hidroxiecdizona (20he) și hormonul juvenil III (JH3) în ouă diapausing și non-diapausing la trei puncte de timp care paranteză etapele cheie ale dezvoltării embrionare și au interpretat rezultatele noastre în contextul rnaseq colectat anterior (Poelchau și colab.,2013a, b) și date manipulative (Suman și colab., 2015).
materiale și metode
colectarea ouălor
toate măsurătorile au folosit ouă F11 dintr-o colonie de laborator a Ae. albopictus a fost înființat în August 2015 din peste 200 de larve colectate în Manassas, VA, Statele Unite. Înainte de acest experiment, populația a fost menținută în condițiile descrise anterior . Pentru a genera probe de ou pentru acest experiment, larvele F10 au fost menținute în recipiente de Sterlit de 5,5 L la o densitate de aproximativ 250 larve pe 2.5 L apă deionizată (DI) cu 5 mL de suspensie alimentară (Armbruster și Conn, 2006) sub fotoperioadă LD la 21 C și 80% rh. În fiecare luni–miercuri–vineri, larvele au fost filtrate printr-o plasă cu plasă fină, apoi transferate într-un recipient curat de Sterlit de 5,5 L; pupele au fost distribuite în cuști adulte menținute fie sub diapause-evitarea fotoperioadei LD, fie sub diapause-inducerea, fotoperioada de zi scurtă (SD, 8:16 L:D).
femelelor adulte li s-au furnizat săptămânal sânge și o cană de ovipoziție căptușită cu prosop de hârtie nealbită și umplută pe jumătate cu apă DI până la încetarea ovipoziției. Ouăle au fost colectate zilnic, menținute pe un prosop de hârtie umed timp de 48 de ore, apoi uscate la aer și depozitate în recipiente în condiții SD. La 5, 7 și 11 zile post-ovipoziție (dpov), ouăle colectate au fost cântărite până la cel mai apropiat 1 ectgg (medie: 11,460 mg, interval: 9,1–19,5 mg) pe un microbalance MX5 (Mettler-Toledo, Columbia, OH, Statele Unite), plasate în tuburi de 1,5 mL, înghețate în azot lichid între orele 11 am și 12 pm (timp Zeitgeber 3-4 ore) și depozitate la -80 C. Un subset de ouă a fost păstrat pentru a confirma că condițiile de fotoperioadă LD și SD au stimulat producția de ouă non-diapause și respectiv diapause (vezi Urbanski și colab., 2012).
am prelevat ouă la 5, 7 și 11 dpov din două motive. În Primul Rând, Ae. albopictus inițiază diapause (sensu Koutottottotl, 2006; Kouttottottotl și colab., 2017) După finalizarea dezvoltării embrionare (Mori și colab., 1981). În condițiile utilizate în experimentele noastre, Ae. embrionii albopictus suferă segmentare la aproximativ 5 dpov, iar embrionii non-diapause sunt competenți să eclozeze la aproximativ 7 dpov (Poelchau și colab., 2013a). Până la 11 dpov, arestarea dezvoltării este ferm stabilită în embrionii diapause (Poelchau și colab., 2013b). Astfel, am ales să comparăm embrionii diapause și non-diapause la momentele de timp care cuprind perioada de competență pentru incubație, inclusiv stabilirea arestării dezvoltării în timpul diapauzei. În al doilea rând, am ales puncte de timp care au fost similare cu cele utilizate pentru experimentele anterioare RNAseq de embrioni (Poelchau și colab., 2013a, b) astfel încât aceste date să poată fi utilizate pentru a interpreta rezultatele noastre actuale. Aceste rezultate RNAseq s-au dovedit anterior a fi bine corelate cu rezultatele expresiei genelor obținute prin qRT-PCR (Poelchau și colab., 2013a).
ecdysteroid cuantificare
pentru a extrage ecdysteroids, probele de ou au fost plasate într-un tub Borosilicat de sticlă 0.5 mL conic fund micro țesut polizor tub (Wheaton, Millville, NJ, Statele Unite ale Americii), cu 1 hectolitru de farnesol (Sigma-Aldrich, St.Louis, MO, Statele Unite ale Americii) adăugat ca un standard intern. Ouăle au fost omogenizate manual de trei ori cu un pistil de Teflon în MeOH refrigerat de 300 de centi HPLC, apoi filate la 12.000 RCF timp de 10 min într-o centrifugă frigorifică (4 centi c). Supernatantul rezultat a fost adunat apoi eluat prin coloane de oxid de aluminiu de două ori cu meoh răcit 90% și o dată cu MeOH. Probele au fost uscate într-o centrifugă de vid refrigerată (4% c), omogenizată în 20% meoh și depozitată la -80% C până la analiză. Controalele pozitive au fost pregătite prin adăugarea a 1 hectolitru de 20he (Sigma-Aldrich) și farnesol la tuburile goale care altfel au fost prelucrate identic. Analiza eșantionului de concentrație ecdisteroidă a fost efectuată utilizând un Micromass Quattro Micro LC-MS/MS (Waters Co., Milford, MA, Statele Unite). Volumele de injecție au fost de 5,0 unqql, cu separări efectuate folosind coloane ACQUITY UPLC BEH C18 de 2,1 mm pe 50 mm cu o fază staționară de 1,7 unqqm (Waters Co.). Condițiile de funcționare pentru cursele LC au fost un debit de fază mobilă de 0,37 mL min–1 cu o fază mobilă binară de 0,1% acid formic în acetonitril și 0,1% acid formic în apă. Condițiile inițiale au fost 1: 99 acetonitril: apă, urmat de fluxul izocratic timp de 0,3 min. La 0,3 min, un gradient liniar de la 1:99 la 99: 1 acetonitril: apa a fost aplicată peste 4,2 min, urmată de 1,0 min flux izocratic la 99:1 acetonitril:apă, după care faza mobilă a revenit la 1:99 acetonitril:apă. Setările spectrometrului de masă au fost pozitive prin electrospray, cu o temperatură de desolvare de 350 ctx20he și farnesol eluate la 2,89 și respectiv 4,97 min, și au fost cuantificate utilizând monitorizarea reacțiilor multiple a tranzițiilor caracteristice: 481,22 (m/z) > 445,24 (M/z), respectiv 205,41 (m/z) > 121,09 (m/z). Limita de detecție a testului este de aproximativ 5 pg.
cuantificarea hormonului Juvenil
fiecare probă de ou a fost plasată într-un tub Borosilicat de sticlă cu fund conic de 0,5 mL de râșniță micro-țesut (Wheaton) cu 1 hectar de farnesol (Sigma-Aldrich) adăugat ca standard intern. Ouăle au fost omogenizate manual de trei ori cu un pistil de Teflon în hexan refrigerat de 500 de centi HPLC, apoi filate la 12.000 RCF timp de 10 min într-o centrifugă frigorifică (4 centi c). Fracțiunile hexanice au fost recombinate într-un flacon din sticlă borosilicată curată și uscate prin centrifugare în vid. JH a fost cuantificat folosind cromatografia de gaze / spectrometria de masă așa cum s-a descris anterior (Brent și Vargo, 2003). Pe scurt, reziduul a fost spălat din flacoane cu trei clătiri de hexan și adăugat la coloanele de sticlă borosilicată umplute cu oxid de aluminiu. Pentru a filtra contaminanții, probele au fost eluate prin coloane succesiv cu hexan, 10% eter etilic-hexan și 30% eter etilic-hexan. După uscare, probele au fost derivatizate prin încălzire la 60 centimetric C timp de 20 min într-o soluție de alcool metil-D (Sigma-Aldrich) și acid trifluoroacetic (Sigma-Aldrich). Probele au fost uscate, omogenizate în hexan și eluate din nou prin coloane de oxid de aluminiu. Componentele nededicatizate au fost îndepărtate cu 30% eter etilic. Derivatul JH a fost colectat în flacoane noi prin adăugarea a 50% acetat de etil-hexan. După uscare, probele au fost resuspendate în hexan, apoi analizate folosind o serie HP 7890a GC (Agilent Technologies, Santa Clara, CA, Statele Unite) echipată cu o coloană de ceară Zebron ZB de 30 m 0,25 mm (Phenomenex, Torrance, CA, Statele Unite) cuplată la un detector selectiv de masă inertă HP 5975c. Heliul a fost folosit ca gaz purtător. Forma JH a fost confirmată prin primele probe de testare în modul de scanare pentru semnăturile cunoscute de JH 0, JH I, JH II, JH3 și JH3 etil; JH3 a fost confirmat ca formă endogenă primară la această specie. Probele ulterioare au fost analizate folosind modul MS SIM, monitorizând la m / z 76 și 225 pentru a asigura specificitatea derivatului d3-metoxihidrină al JH3. Abundența totală a fost cuantificată în raport cu o curbă standard a derivatului JH3 și ajustată pentru masa inițială a ouălor. Limita de detecție a testului este de aproximativ 1 pg.
analize statistice
analizele au fost efectuate în R (R Core Team, 2019). Abundența JH3 a fost transformată în jurnal și analizată folosind ANOVA pentru a testa efectele statutului diapauzei, vârstei și interacțiunii lor. 20el abundența a fost neliniară și potrivită folosind două modele de regresie polinomială: un model complet care include vârsta și starea de dezvoltare (diapause, non-diapause) și un model redus care conține doar vârsta. Modelele au fost comparate folosind ANOVA pentru a determina dacă includerea stării de dezvoltare a îmbunătățit semnificativ puterea explicativă.
analizele transcripționale au utilizat datele rnaseq colectate anterior de la diapause și non-diapause ae. ouă albopictus (set suplimentar de date 1; Poelchau și colab., 2013a, b). Genele exprimate diferențiat la 3, 6 și 11 dpov au fost investigate prin analize de îmbogățire a setului de gene (Mootha și colab., 2003) folosind pachetul de pian (V Otrivtremo și colab., 2013) setați la parametrii impliciți urmați de corecția FDR. Un set de gene de referință a fost adnotat cu Termenii biologici process Go atribuiți pe VectorBase (Giraldo-Calder XVN și colab., 2015). Genele căii de sinteză JH3 și ecdisteroide, precum și receptorii JH3 și genele de degradare au fost adnotate manual conform literaturii anterioare (Bai și colab., 2007; Li și colab., 2011; Nouzova și colab., 2011; Niwa și Niwa, 2014; Matthews și colab., 2016). În plus, am adnotat genele JH3 și ecdysteroid-inductibile pe baza descrierilor genice furnizate în Vectorbază (Giraldo-Calder XVN și colab., 2015). O prezentare completă a acestor seturi de gene este furnizată în setul suplimentar de date 1.
rezultate
incidența medie a diapauzei a fost de 95,9% (interval: 90,5–100%) sub fotoperioada SD și 5.4% (Interval: 1,8–9,5%) sub fotoperioada LD (set suplimentar de date 2); astfel, tratamentele noastre fotoperiodice au indus răspunsurile diapause așteptate. 20EL a atins punctul culminant atât în condiții diapause, cât și în condiții non-diapause la 7 dpov,dar includerea statutului diapause ca factor într-un model pătratic pentru 20he abundența nu a îmbunătățit puterea explicativă a modelului (figura 1a și setul de date suplimentar 3; F3, 43 = 0,21, p = 0,890). Aceste rezultate indică faptul că statusul diapauzei nu explică în mod semnificativ variația titrului 20HE. Analiza căii RNAseq a indicat că nici calea de sinteză 20HE, nici genele inductibile 20he nu au fost modificate semnificativ în condiții diapause vs. non-diapause la 3, 6 sau 11 dpov (Tabelul 1).

Tabelul 1. Rezultatele analizei îmbogățirii seturilor de Gene, a se vedea setul de date suplimentar 1 pentru un rezumat complet al acestor seturi de gene.
JH3 a fost confirmat a fi forma primară a hormonului juvenil la această specie. Abundența JH3 a fost redusă semnificativ la embrionii diapause (figura 1b și setul suplimentar de date 3; F1,64 = 5,08, p = 0,028). Cu toate acestea,vârsta (F2, 64 = 0,66,p = 0,518) și interacțiunea statusului diapauzei și a vârstei (F2, 64 = 0,13, p = 0,883) nu au afectat nivelurile de JH3. Analiza căii a datelor RNAseq a indicat că calea de sinteză a JH3 a fost reprimată semnificativ la 3 dpov și genele inductibile JH au fost reprimate semnificativ la 6 dpov (Figura 2 și Tabelul 1).
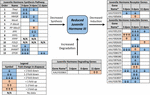
Figura 2. Rezumatul expresiei diferențiale a genelor implicate în sinteza, degradarea și inducția JH3 pentru Ae. albopictus (Poelchau și colab., 2013a, b). Reducerile semnificative induse de diapauză (albastru) sau creșteri (portocaliu) sunt indicate de celulele umbrite. Magnitudinea diferențială este indicată de săgeți așa cum este descris în legenda din stânga jos. Genele fără nume comune sunt identificate prin numărul de identificare al Vectorbazei.
discuție
Diapauza este o adaptare pe scară largă care permite insectelor să-și alinieze creșterea și reproducerea cu condiții favorabile sezonului (Tauber și colab., 1986; Danks, 1987). Înțelegerea modificărilor hormonale prin care insectele coordonează percepția indiciilor externe, care induc diapause, cu mecanismele fiziologice care duc la stoparea dezvoltării este un obiectiv de lungă durată în biologie (Lees, 1956; Denlinger, 2002). Decenii de cercetare au condus la identificarea strategiilor hormonale comune pentru diapauză la etapele larvare, pupale și adulte; cu toate acestea, reglarea hormonală a diapauzei embrionare rămâne în mare parte necunoscută, în special în Diptera (Denlinger, 1985, 2002; Denlinger și colab., 2012). În acest studiu, măsurarea noastră directă a abundenței reduse a JH3 în diapauză este coroborată de profilarea anterioară a expresiei genelor (Poelchau și colab., 2013a, b) și experimente manipulative (Suman și colab., 2015) susținând ipoteza că JH3 reglementează diapauza embrionară la această specie.
Ecdysteroids
în studiul de față, am măsurat direct abundența ecdysteroid la trei puncte de timp în condiții diapause și non-diapause la embrionii țânțarului ae. albopictus. În concordanță cu rezultatele anterioare ale Toxorhynchites amboinensis (Russo și Westbrook, 1986), nivelurile 20HE au atins punctul culminant aproape de încheierea dezvoltării embrionare (7 dpov; figura 1a). Cu toate acestea, titrul 20HE nu a diferit semnificativ între diapause și ouăle non-diapause. Mai mult, nici calea de sinteză 20HE, nici genele inductibile 20HE nu au suferit modificări semnificative, coordonate în expresia genelor (Tabelul 1). Concluzionăm că, spre deosebire de diapauzele embrionare examinate anterior în Lepidoptera și Ortoptera (Hasegawa, 1963; Yamashita și Hasegawa, 1966; Gregg și colab., 1987; Yamashita, 1996; Lee și Denlinger, 1997; Lee și colab., 1997; Tawfik și colab., 2002), 20EL nu reglementează inițierea diapauzei în Ae. albopictus.
hormonul Juvenil
în contrast, am observat o abundență de JH3 de aproximativ două ori mai mică în embrionii diapause (figura 1b). În timp ce magnitudinea este mai mică decât diferențele în abundența JH3 detectate în condiții diapause vs.non-diapause la insecte care diapause în alte etape ale vieții (de exemplu, Yin și Chippendale, 1979; Walker și Denlinger, 1980; Readio și colab., 1999), măsurătorile noastre directe sunt coroborate cu dovezi transcripționale (Figura 2). De exemplu, calea de sinteză JH3 este reprimată semnificativ în timpul preparării timpurii a diapauzei (3 dpov). Susținând interpretarea noastră a acestor rezultate, studiile anterioare au descoperit că expresia redusă a genelor de-a lungul căii de sinteză a JH3 este puternic corelată cu abundența JH3 mai mică în ambele Aedes aegypti (Nouzova și colab., 2011; Rivera-Perez și colab., 2014) și Diploptera punctata (Huang J. și colab., 2015). Genele inductibile JH sunt, de asemenea, reprimate semnificativ în timpul preparării diapauzei târzii (6 dpov; Tabelul 1), sugerând că embrionii destinați diapauzei au o capacitate limitată de a răspunde la semnalizarea JH3 aproape de momentul în care embrionii non-diapause devin competenți să eclozeze. Mai mult, expresia genelor pentru o enzimă degradantă JH3 (esteraza JH) este reglată în mod semnificativ în întreținerea diapauzei timpurii (11 dpov; Figura 2), precum și mai târziu în întreținerea diapauzei (21 și 40 dpov; Poelchau și colab., 2013b) sugerând că JH3 poate rămâne la abundență scăzută pe tot parcursul diapauzei la această specie.
abundența redusă a JH3 în timpul stopului de dezvoltare este oarecum contraintuitivă, deoarece titrul scăzut de hormoni juvenili este de obicei asociat cu mutări progresive, în special de la stadiul larvar la pupal (Palli, 2016). Cu toate acestea, aplicarea unui analog JH3 (pyriproxyfen) la diapause Ae. ouăle albopictus accelerează rata de întrerupere a diapauzei într-o manieră dependentă de doză (Suman și colab., 2015). La cea mai eficientă doză, aproximativ 80% din ouăle expuse la piriproxifen termină diapauza cu 30 dpov, comparativ cu doar 1% încetarea diapauzei cu 80 dpov pentru ouăle de control (Suman și colab., 2015). Modificările abundenței JH3 pot genera, de asemenea, un fenotip asemănător diapauzei embrionare la alte specii. De exemplu, în B. mori, reducerea experimentală a sintezei JH3 de către jhamt knockout are ca rezultat larve farate complet dezvoltate care nu răspund la stimulii de incubație; acest fenotip poate fi salvat într-o manieră dependentă de doză prin aplicarea fie a JH3 extras, fie a metoprenului, a analog JH3 (Nakao și colab., 2015). În mod similar, suplimentarea experimentală a unui alt analog al hormonului juvenil (RO-20-3600) la ouăle melanogaster Drosophila nou depuse produce larve de farate care sunt mobile în ou, dar nu reușesc să eclozeze (Smith și Arking, 1975). Împreună, aceste date indică faptul că abundența hormonilor juvenili contribuie probabil la reglarea răspunsurilor de incubație la diverse insecte. Diapauza embrionară în stadiul larvar farat este definită de un eșec de a răspunde la stimulii de incubație astfel, modificările abundenței hormonilor juvenili pot reprezenta o strategie hormonală comună pentru reglarea acestei forme de diapauză.
în concordanță cu datele transcripționale și manipulative anterioare, rezultatele noastre implică puternic abundența redusă a JH3 ca regulator probabil al diapauzei embrionare în Ae. albopictus. Această cercetare este prima cuantificare directă a abundenței hormonale în timpul diapauzei embrionare la orice specie Dipterană și reprezintă un pas important spre clarificarea reglării hormonale a acestei adaptări cruciale la mediile adverse. Caracterizarea legăturii dintre stimulii externi și stoparea dezvoltării prin semnalizarea hormonală rămâne o provocare majoră (Denlinger, 1985; Denlinger și colab., 2012). Elucidarea acestor căi într-o varietate de specii va oferi o perspectivă critică pentru problemele fundamentale și aplicate, inclusiv înțelegerea bazei moleculare și fiziologice a evoluției istoriei vieții, prezicerea răspunsurilor speciilor la schimbările climatice și identificarea unor ținte noi pentru gestionarea speciilor dăunătoare și vectoriale (Denlinger, 2008).
Declarație privind disponibilitatea datelor
toate seturile de date generate pentru acest studiu sunt incluse în articol/Material suplimentar.
contribuțiile autorului
ZB, CB și PA au contribuit la conceperea, proiectarea studiului și au pregătit proiectul inițial al manuscrisului. ZB, MM și JS au efectuat creșterea animalelor și au colectat probele de ouă. CB a efectuat analizele LC-MS / MS și GC-MS. ZB a efectuat analizele statistice. Toți autorii au contribuit la revizuirea manuscrisului și au aprobat versiunea trimisă.
finanțare
această lucrare a fost susținută de Institutele Naționale de sănătate (1r15a|111328-01) și dotarea familiei Davis către PA.
Conflict de interese
autorii declară că cercetarea a fost realizată în absența oricăror relații comerciale sau financiare care ar putea fi interpretate ca un potențial conflict de interese.
mulțumiri
mulțumim lui Allan Knopf și Dan Langhorst pentru asistență în măsurarea titrurilor hormonale. De asemenea, Îi mulțumim lui David Denlinger pentru comentariile utile cu privire la versiunile anterioare ale manuscrisului. Utilizarea denumirilor comerciale sau a produselor comerciale din acest articol este exclusiv în scopul furnizării de informații specifice și nu implică recomandări sau aprobări din partea Departamentului Agriculturii al Statelor Unite. USDA este un furnizor de șanse egale și angajator.
Material Suplimentar
Echipa De Bază R (2019). R: un limbaj și un mediu pentru calculul statistic. Viena: Echipa De Bază R.
Google Scholar
V Otrivtremo, L., Nielsen, J. și Nookaew, I. (2013). Îmbogățirea analizei setului de gene a datelor la nivel de genom prin încorporarea direcționalității expresiei genelor și combinarea ipotezelor și metodelor statistice. Acizi Nucleici Res. 41, 4378-4391. doi: 10.1093/nar / gkt111
PubMed Abstract / CrossRef Full Text / Google Scholar