introduktion
L1-celladhesionsmolekylgenen (L1CAM) är en neuronal celladhesionsmolekyl som tillhör immunoglobulinens superfamilj; den har nyckelfunktioner i utvecklingen av nervsystemet (Itoh och Fushiki, 2015). Mutationer i L1CAM har varit relaterade till X-länkade neurologiska syndrom, som sammanfattas som L1-sjukdomar. De klassificeras enligt följande: X-länkad hydrocephalus (XLH) på grund av stenos av akvedukten av Sylvius (HSAS), MASA syndrom (intellektuell funktionsnedsättning, afasi, shuffling gång, adducted tummar), spastisk paraparesis typ 1 (SP1) och X-länkad agenes av corpus callosum (ACC) (Weller och Gartner, 2001; Itoh och Fushiki, 2015).
cirka 282 sjukdomsframkallande mutationer (DMs) i l1cam-genen har rapporterats i hgmd bisexuell Professional 2019.2 (https://portal.biobase-international.com/hgmd/pro/all.php). Förändringar i l1cam-genen varierar; mutationsdataanalyser från 282 patienter avslöjar 51% missense-och nonsensmutationer, 25% deletioner, 5% Infogningar och 19% skarvplatsändringar, men tysta mutationer i L1CAM med patogen potential var sällsynta, och tysta mutationer ignorerades ofta särskilt i Wes-detektion (Wes).
i denna studie, med hjälp av WES, screenade vi fetalt DNA hos en kinesisk gravid kvinna som har rapporterat fem kontinuerliga graviditeter med fosterhydrocephalus; vi hittade bara en ny tyst mutation c.453g > T (p.Gly151 = ) i l1cam-genen. Intressant, genom ytterligare analys, indikerade vi den tysta mutationen skapade en potentiell 5 ’ skarvplats konsensussekvens, vilket skulle resultera i en in-frame-radering av 72 bp från exon 5 och 24 aminosyror av l1cam-proteinet.
Fallpresentation
en 28-årig frisk kvinna hänvisades till vår klinik efter fyra frivilliga avslutningar av graviditeten på grund av fosterhydrocefalus på andra sjukhus. Alla Foster var män. När hon anlände till vårt sjukhus (Women ’ s Hospital, School of Medicine, Zhejiang University, Zhejiang, Kina) var hon redan på sin femte graviditet vid 24 veckors graviditet, med en fosterhydrocefalus genom bildundersökningar. För att utforska den genetiska orsaken genomfördes fosterblodprovtagning vid 26 veckors graviditetsålder. Konventionella cytogenetiska studier utfördes för både foster-och föräldraprover, och fosterprovet analyserades vidare med enkelnukleotidpolymorfism (SNP) array och WES.
denna studie genomfördes i enlighet med rekommendationerna från Etikutskottet för kvinnors sjukhus, School of Medicine Zhejiang University, och informerat samtycke förvärvades från alla deltagare i denna studie i enlighet med Helsingforsdeklarationen. Studieprotokollet godkändes av Granskningsnämnden för Women ’ s Hospital, School of Medicine, Zhejiang University i Kina.
material och metoder
karyotyp och SNP-Array
karyotyperna av fetalt navelsträngsblod och perifert navelsträngsblod bestämdes genom konventionell karyotypning av minst 30 blodlymfocyter, som arresterades vid metafas av kolchiciner. G-banding karyotyper av odlade celler utfördes på 320-400-bandnivå med en upplösning på cirka 10 Mb. SNP-array utfördes av CytoScan 2600000 markörer inklusive 750 000 SNP-prober och 1 900 000 icke-polymorfismprober för omfattande helgenomtäckning. Data analyserades av kromosom Analysis Suite (ChAS) programvara (Affymetrix, Santa Clara, CA) baserat på grch37/hg19-enheten. Rapporteringströskeln för resultatet av kopianummer fastställdes till 500 kb med ett markörantal på 50 för vinster och till 200 kb med ett markörantal på 50 för förluster.
hel-Exome sekvensering
huvuddelen av WES tillhandahölls av Beijing Genomics Institute. Genomiskt DNA extraherades av ett Dneasy-Blodkit (Qiagen, CA) och fragmenterades sedan av Covaris LE220 (Massachusetts, USA) för att generera ett parat bibliotek (200-250 bp). Alla förstärkta bibliotek utfördes på bgiseq-500-plattformen, ENKELSTRÄNGSDNAET blandades med mgieasy bisexuell DNA Library Prep Kit V1 (BGI, Shenzhen, Kina) och sekvenserades sedan med 100SR-kemi med bgiseq-500Rs sekvenseringssats med hög genomströmning (BGI, Shenzhen, Kina).
Clean läser (med en längd av 90 bp) härledd från riktad sekvensering och filtrering anpassades sedan till human genome reference (hg19) med hjälp av Burrows-Wheeler Aligner (BWA) Multi-Vision programpaket (Li och Durbin, 2009). Efter justering användes utdatafilerna för att utföra sekvenseringstäckning och djupanalys av målregionen, enkelnukleotidvarianter (SNV) och indel calling, vi använde GATK-programvaran för att upptäcka SNV och indels (McKenna et al., 2010), alla SNV och indels filtrerades och uppskattades via flera databaser, inklusive National Center for Biotechnology Information (NCBI) Single-Nucleotide Polymorphism Database (dbSNP), HapMap, 1000 Genome Project dataset och databas med 100 Kinesiska friska vuxna. Vi använde Condel, SIFT, PolyPhen-2, LRT, Mutation Taster och PhyloP för att förutsäga effekten av varianter. Patogena varianter bedöms enligt protokollet utfärdat av American College of Medical Genetics and Genomics (ACMG) (Richards et al., 2015). Human gen Mutation Database (Hgmd) användes för att screena mutationer. Alla potentiella patogena varianter validerades med Sanger-sekvenseringsmetoder.
RNA-extraktion, PCR och sekvensering
mononukleära celler i perifert blod (Pmbcs) och mononukleära celler i navelsträngsblod (CBMC) isolerades genom Ficoll-täthetsgradientseparation. Totalt RNA extraherades från PMBCs och CBMCs med användning av TRIzol (Takara, Japan). Extraherade totala rna transkriberades omvänd med RT Kit (Takara, Japan). PCR utfördes med GoldStar Best Master Mix (Cwbio, Peking). Primersekvenser listas: L1CAM-DNA-5F, CCCACCCGTCCTTTCCTA; L1CAM-DNA-5R, CGCTCGTCCTGCTTGATGT; L1CAM-mRNA-4-6-F, GGTGTCCACTTCAAACCCAA; och L1CAM-mRNA-4-6-R, GCGGCTTCCTGTCAATCA. Sanger-sekvensering utfördes av en ABI 3130 DNA-analysator.
resultat
en 28-årig frisk kvinna hänvisades till vår klinik efter fyra frivilliga avslutningar av graviditeten på grund av fosterhydrocefalus. Alla Foster var manliga (Figur 1A). Den familjära stamtavlan tycktes visa XLH. Hon var redan här femte graviditeten vid 26 veckors graviditet. Fetal ventrikulomegali detekterades genom fetal ultraljudsskanning och MR, vilket konsekvent visade närvaron av hydrocephalus. De visade att den bilaterala hjärnkammaren och den tredje ventrikeln uppenbarligen utvidgades, och det fanns svår hydrocephalus i intracerebral och agenes av corpus callosum (Figur 1b).
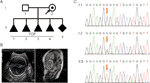
Figur 1 (A) stamtavla i familjen. TOP, uppsägning av graviditet. (B) avbildningsundersökningar av fostret. Fetal ultraljudsundersökning och fetal MRI visade att det fanns svår hydrocephalus i fostret. (C) sekvensanalys av genomiskt DNA från familjemedlemmar. Genotyperna för L1CAM var vildtyp, c. 453g > t Het och c. 453G > t Hom, i I:1 (make), I:2 (gravid kvinna) och II:5 (Foster). Mutationen indikeras av de röda pilarna.
för att undersöka den möjliga genetiska orsaken utförde vi karyotypanalys och SNP-array för att analysera fostrets blodprovtagning och fann inga positiva resultat. Koroidal neovaskularisering (CNV) resultat har deponerats i genuttrycket Omnibus (GEO); anslutningsnumret är GSE133063, som bifogas nedan (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE133063).
sedan upptäckte vi fostret av WES. Den analytiska strategin för att hitta sannolik patogen variantidentifiering visades i Figur S1. En lista över varianter (tabell S1) erhölls genom screening av variantfrekvenser, mutationsstatus och arvläge. Med hänsyn till de hydrocephalusassocierade generna (HP:0000238, http://compbio.charite.de/hpoweb/showterm?id=HP:0000238#id=HP_0000238) (tabell S2) fanns det ingen ytterligare anmärkningsvärd mutation förutom den tysta mutationen av c.453g > T i exon 5 av l1cam-genen (NM_000425.3). c.453G > t rapporterades inte i HGMD och ClinVar och hittades inte i dbSNP, gnomAD och andra dataset. Enligt ACMG: s standarder och riktlinjer (Richards et al., 2015), hade det ännu inte nått kriteriet ”patogen” eller ”sannolikt patogen”, men det fanns inga andra potentiella mutationer; vi hade inget annat val än att göra en ytterligare analys av den tysta mutationen som hittades.
enligt traditionellt tänkande inträffade denna bassubstitution i den tredje basen i kodon 151, som kodar för en glycin och därigenom skapar en neutral mutation (s.Gly151 = ). Denna variant bekräftades i DNA extraherat från fetalt navelsträngsblod och perifert blod i paret genom Sanger-sekvensering (figur 1C). Kvinnan bar den heterozygota mutationen, och hennes man var en vildtypsgenotyp.
med Mutationsprovning (http://www.mutationtaster.org/), C.453g > t fick poäng som ”sjukdom som orsakar.”Det visade att proteinfunktioner kan påverkas och skarvplatsen kan ändras; vi var nyfiken på de potentiella skarvningseffekterna av l1cam-funktionen i denna tysta mutation. Den tysta mutationen testades med hjälp av följande online-mjukvaruprodukter: NetGene2 (http://www.cbs.dtu.dk/services/NetGene2/) och NNSplice (http://www.fruitfly.org/seq_tools/splice.html); 5′ – potentiella skarvplatsen förutspåddes också att skapas i L1CAM c.453g > t-mutation med hjälp av programvaruprodukterna (figur S2). Resultaten visade att denna tysta mutation skapade en potentiell 5′ skarvplats 72 bp uppströms från den normala exon 6/intron 6 skarvplats (figur 2a). Om så är fallet kan vi hitta längdförändringen av l1cam messenger RNA (mRNA) mellan I:2 och II:5 (figur 2a). RT-PCR utfördes med användning av primers utformade för att förstärka exoner 4-6 i l1cam mRNA. Resultaten visade faktiskt ett kort band av trunkering i fetal cDNA PCR (II: 5), medan bandet förstärkt från l1cam mRNA innehöll det förväntade långa bandet i make cDNA PCR (I:1) och långa/korta band i den gravida kvinnan cDNA PCR (i:2) (Figur 2B). Direkt sekvensering av det förstärkta fragmentet visade att deletionen involverade den sista 72 bp av exon 5 i manlig fetal cDNA (kvinnan var bärare) (figur 2C). Vi har de avgörande patogena bevisen.
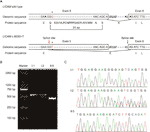
Figur 2 (A) Schematisk representation av exon 5, intron 6 och exon 6 organisation i L1CAM. (B) RT-PCR-analys av exoner 5 och 6 av l1cam cDNA från perifera mononukleära blodceller (PMBCs) och mononukleära celler i navelsträngsblod (CBMC). Agarosgelelektrofores av RT-PCR-produkter genererade från I:1 (make), I:2 (gravid kvinna) och II:5 (Foster). (C) sekvensanalys av RT-PCR-produkten från pmbcs hos paret och CBMC hos fostret.
denna tysta mutation resulterade i 24 aminosyror av l1cam-protein (rester 151-174); Lys (K) ersattes av Glu (E) vid kodon 175 (figur 2a). Det fanns anpassning av flera l1cam-proteinsekvenser över flera arter och bevarande av de saknade aminosyrorna i L1CAM över däggdjur: Homo sapiens, Pan troglodytes, BIM taurus, Mus musculus och Rattus norvegicus (figur 3A). Vildtyp och c. 453G > t-skarvmutation L1CAM-proteiner förutspåddes av programvaran CPHmodels-3.2 Server (http://www.cbs.dtu.dk/services/CPHmodels/) (figur 3b). Immunoglobulinliknande (Ig-liknande) domän 2 (rester 134-230) av vildtyp och skarvmutation L1CAM-proteiner visas i Figur 3C. L1CAM c. 453G > t skarvmutation förändrade proteinstrukturen, särskilt den Ig-liknande domänen 2
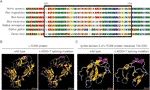
Figur 3 (A) anpassning av flera l1cam-proteinsekvenser över arter. L1cam c.453G > t resulterade i att 24 aminosyror av l1cam-protein (rester 151-174) saknades i den konserverade aminosyraregionen i olika arter. Den svarta kolumnen visar de saknade aminosyrorna. (B) strukturerna av vildtyp och c.453g > t Splitsning mutation L1CAM protein som förutsagts av programvaran CPHmodels-3.2 Server. (C) strukturerna för immunoglobulinliknande (Ig-liknande) domän 2 (rester 134-230) av vildtyp och skarvmutation L1CAM-protein.
diskussion
tysta mutationer upptäcktes ofta av WES, men otillräcklig uppmärksamhet har betalats, vilket leder till utelämnande av DMs. I denna studie använde vi WES för att utforska den genetiska orsaken till en kinesisk familj med hydrocephalus men hittade bara en ny tyst mutation i L1CAM, vilket tvingade oss att göra en ytterligare analys. Lyckligtvis visade vi att den tysta mutationen skapade en ny 5′ skarvplats och var en DM.
mutationer i L1CAM kan orsaka en X-länkad L1-sjukdom, men kliniska symtom är varierande; mutationer producerar oväntade fenotyper. I studien är de fem lidande fostren alla män, vilket överensstämmer med ett arvsmönster. Fostrets ultraljudsskanning och MR visar en typisk L1-sjukdom, inklusive XLH och agenes av corpus callosum. Det förbättrar vår förståelse för genotyp-fenotyp korrelation av L1CAM.
L1CAM c. 453G > T (p.Gly151 = ) ansågs ursprungligen inte ha någon effekt på proteinsekvensen. Men andra tysta mutationer, c. 924C > T (p.Gly308 = ) och c.645c > T (p.Gly215 = ), i l1cam-genen har rapporterats vara DMs (Du et al., 1998; Vos et al., 2010). C.924c > t-mutation resulterade i aktivering av en ny skarvplats 69 bp 5′ till det normala exon 8/intron 8 donatorskarvplats, och det har förklarats som en ”sjukdomsframkallande” plats för hydrocephalus (Du et al., 1998). För c.645c > T i L1CAM raderades 51 bp med aktiveringen av en ny exon 6/intron 6 donor splice (Vos et al., 2010). Vår nuvarande studie var liknande; mutationen av c.453g > t skapade en potentiell 5′ skarvplats uppströms från den normala exon 5/intron 5 skarvplats. Alla dessa tysta mutationer skapade nya donatorskarvplatser, vilket resulterade i att exonen hoppades över. Det påminde oss om att ägna stor uppmärksamhet åt dessa tysta mutationer, vilket kan påverka skarvning av proteiner.
som ett transmembrant glykoprotein och en medlem av immunoglobulinsuperfamiljen av celladhesionsmolekyler kan l1cam-proteinet interagera på cellytan med ett antal olika glykoproteiner, och homofil bindning är förmodligen dess huvudsakliga interaktionssätt (Wei och Ryu, 2012). Studierna på kristallstrukturen hos IG-liknande domäner 1-4 i neurofascin föreslog att många patologiska L1-mutationer påverkar konserverade aminosyrarester inom dessa domäner och stör homofila interaktioner (Liu et al., 2011), särskilt som verifierats av funktionsforskningen av IG-liknande domän 2 (Zhao et al., 1998). I vår studie spekulerade vi i att L1CAM c.453g > t förändrade ig-liknande domän 2 i den extracellulära delen av l1cam-proteinet, vilket ledde till den onormala extracellulära interaktionen, utan att börja initiera nedströms signalvägen. En ytterligare studie tillägnad masspektrometri av denna l1cam-variant skulle klargöra specifikt vilket molekylärt ensemble som produceras i cellen.
Sammanfattningsvis rapporterade vi genom WES en ny tyst mutation c.453g > T i L1CAM som producerar en 5′ skarvplats ansvarig för hydrocephalus. Denna onormala proteinvariant förutspåddes förändra ig-liknande domän 2, vilket kan påverka l1cam-proteinhomofil bindning. Dessutom utförde vi prenatal genetisk diagnos för den gravida kvinnan som rapporterade fem kontinuerliga graviditeter med hydrocephalus. Under tiden föreslog det att vissa tysta mutationer som detekterades i WES inte skulle ignoreras; splitsningsprognoser av dessa mutationer var nödvändiga. Det gav en ny genetisk grund för prenatal diagnos och preimplantation prenatal diagnos av hydrocephalus.
datatillgänglighet
offentligt tillgängliga dataset analyserades i denna studie. Dessa data finns här: GSE133063 (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE133063).
Etikuttalande
studierna med mänskliga deltagare granskades och godkändes av Granskningsnämnden för Women ’ s Hospital, School of Medicine, Zhejiang University i Kina. Skriftligt informerat samtycke till att delta i denna studie tillhandahölls av deltagarnas vårdnadshavare/anhöriga. Skriftligt informerat samtycke erhölls från den enskilde(s), och mindre(s) förmyndare/anhöriga, för publicering av eventuella identifierbara bilder eller data som ingår i denna artikel.
Författarbidrag
Ys, YLi, MC, YLu, YQ och YY genomförde experiment. YS förberedde siffrorna. MC och YLi analyserade WES-data. YLu och YQ utförde karyotypanalys och SNP-array. YY rekryterade prover. HL och FL tillhandahöll bildundersökningar. YS och MD skrev manuskriptet. Alla författare läste och godkände det slutliga manuskriptet.
finansiering
denna studie stöddes av National Natural Science Foundation of China (Grant No.81801441), det viktigaste forsknings-och utvecklingsprogrammet i Zhejiang-provinsen (Grant No. 2019c03025), Kinas nationella Nyckelforsknings-och utvecklingsprogram (Grant No.2016yfc1000703) och Medical Scientific Research Foundation i Zhejiang-provinsen (Grant No. 2014kya246).
intressekonflikt uttalande
författarna förklarar att forskningen genomfördes i avsaknad av kommersiella eller finansiella relationer som kan tolkas som en potentiell intressekonflikt.
bekräftelser
vi tackar de patienter som är inskrivna i denna forskning. Vi tackar Dr. Jiong Gao (BGI Genomics, BGI-Shenzhen, Shenzhen 518083, Kina) för hans hjälp under utarbetandet av detta manuskript.
tilläggsmaterial
Tilläggsmaterialet för denna artikel finns online på: https://www.frontiersin.org/articles/10.3389/fgene.2019.00817/full#supplementary-material
figur S1 / analytisk strategi för att hitta sannolik patogen variantidentifiering av WES.
figur S2 / Donatorskarvplatser förutsagda av NetGene2 och NNSplice.
tabell S1 / lista över varianter genom screening av varianter frekvenser, mutationsstatus och arv läge.
tabell S2 / lista över hydrocephalus-associerade gener (Export för HP:0000238).
Du, YZ, Dickerson, C., Aylsworth, A. S., Schwartz, Ce (1998). En tyst mutation, C924T (G308G), i l1cam-genen resulterar i X-länkad hydrocephalus (HSAS). J. Med. Genet. 35 (6), 456–462. doi: 10.1136/jmg.35.6.456
PubMed Abstrakt / CrossRef Fulltext / Google Scholar
Itoh, K., Fushiki, S. (2015). Rollen av L1cam i Murin kortikogenes och patogenesen av hydrocephalus. Pathol. Int. 65 (2), 58–66. doi: 10.1111/stift.12245
PubMed Abstrakt / CrossRef Fulltext / Google Scholar
Li, H., Durbin, R. (2009). Snabb och exakt kortläsning med Burrows-Wheeler transform. Bioinformatik 25 (14), 1754-1760. doi: 10.1093 / bioinformatik / btp324
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Liu, H., Focia, P. J., He, X. (2011). Homofil adhesionsmekanism av neurofascin, en medlem av L1-familjen av neurala celladhesionsmolekyler. J. Biol. Chem. 286 (1), 797–805. doi: 10.1074 / jbc.M110.180281
PubMed Abstrakt / CrossRef fulltext / Google Scholar
A., Hanna, M., Banks, E., Sivachenko, A., Cibulskis, K., Kernytsky, A., et al. (2010). Genomanalysverktyget: en MapReduce-ram för analys av nästa generations DNA-sekvenseringsdata. Genom Res. 20 (9), 1297-1303. doi: 10.1101 / gr.107524.110
PubMed Abstrakt / CrossRef fulltext / Google Scholar
han är en av de mest kända och mest kända i världen. (2015). Standarder och riktlinjer för tolkning av sekvensvarianter: en gemensam konsensusrekommendation från American College of Medical Genetics and genomics och Association for Molecular Pathology. Genet. Med. 17 (5), 405–424. doi: 10.1038 / gim.2015.30
PubMed Abstrakt / CrossRef fulltext / Google Scholar
det är en av de mest populära och mest populära. (2010). Genotyp-fenotyp korrelationer i L1 syndrom: en guide för genetisk rådgivning och mutationsanalys. J. Med. Genet. 47 (3), 169–175. doi: 10.1136 / jmg.2009.071688
PubMed Abstrakt / CrossRef Fulltext / Google Scholar
Wei, C. H., Ryu, S. E. (2012). Homofil interaktion av L1-familjen av celladhesionsmolekyler. Exp. Mol. Med. 44 (7), 413–423. doi: 10.3858 / emm.2012.44.7.050
PubMed Abstrakt / CrossRef Fulltext / Google Scholar
Weller, S., Gartner, J. (2001). Genetiska och kliniska aspekter av X-länkad hydrocephalus (L1-sjukdom): mutationer i l1cam-genen. Brum. Mutat. 18 (1), 1–12. doi: 10.1002 / humu.1144
PubMed Abstract | CrossRef Full Text | Google Scholar
Zhao, X., Yip, P. M., Siu, C. H. (1998). Identification of a homophilic binding site in immunoglobulin-like domain 2 of the cell adhesion molecule L1. J. Neurochem. 71 (3), 960–971 doi: 10.1046/j.1471-4159.1998.71030960.x.
PubMed Abstract | CrossRef Full Text | Google Scholar