resultater og diskussion
vi har forstærket 307 BP-fragmentet af mitokondrie-cytokrom B-gener fra de eurasiske oddere Lutra lutra (Letland), L. lutra (Kina), L. lutra (Europa) og L. lutra (asahikava), den asiatiske lille kløede Odder aonyks cinerea og væselene Mustela sibirica og Mustela itatsi, subklonerede de forstærkede produkter i puc18 og bestemte deres sekvenser. Sekvenserne af L. lutra (Europa) og L. lutra (Letland) var helt enige med L lutra (Kina) og L lutra (Letland). Sekvenserne af to individer af Mustela itatsi var identiske, men forskellige i tre positioner end den, der blev rapporteret for nylig (Masuda og Yoshida, 1994), i sekvenspositionerne 1-224 (se Fig. 1).
Fig. 1.
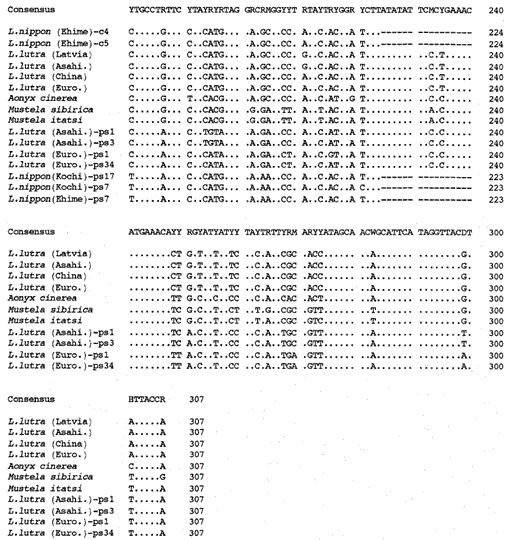
sammenligning af nukleotidsekvenserne af mitokondrie-cytokrom b og cytokrom B-lignende sekvenser af Lutra nippon (Ehime og Kochi) med sekvenserne af L lutra (Letland, Kina, Europa og Asahikava), aonyks cinerea, Mustela sibirica og M. itatsi. Sekvenslængden, 224 bp for L. nippon og 307 bp for de andre. Flere repræsentanter for cytochrome b-lignende sekvenser (ps) er vist. Positioner af deletion og afslutning codon i cytochrom b-lignende sekvenser er bokset. Euroområdet., Europa; Asahi.- Asahikaja.
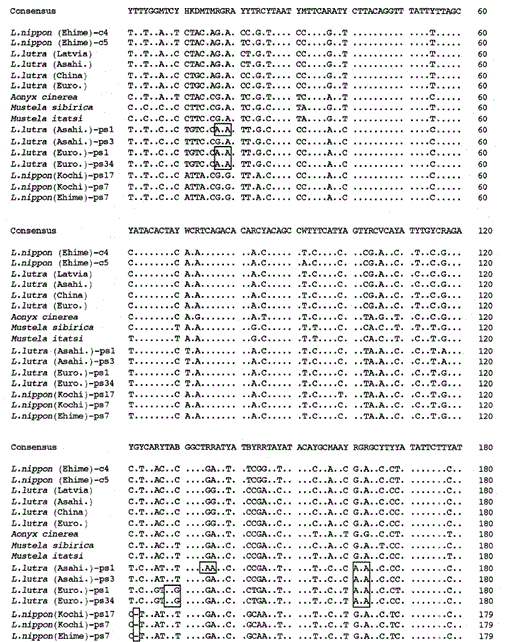
Fig. 1.
fortsat.
på den anden side blev der gjort mange forsøg på at forstærke DNA-fragmentet af cytochrom b-genet af Lutra nippon ved hjælp af forskellige 10-50 år gamle prøver, såsom lever, hjerte og hud bevaret i formalin, det garvede læder og hårene i udstoppet prøve, men alle forsøg var mislykkede.
det er lykkedes os at forstærke 224 bp-fragmentet af mitokondrie cytochrom B-genet fra en 30-årig mumielignende prøve af Lutra nippon fra Ehime (Fig. 1). Direkte sekventering af de forstærkede produkter antydede tilstedeværelsen af mere end to typer sekvens. Derfor blev produkterne subkloneret i pUC18, og 30 kloner blev sekventeret. Blandt dem blev ni kloner tvetydigt identificeret som den mitokondrie cytochrom b-sekvens af Lutra nippon, da sekvensen tilsyneladende lignede mitokondrie-sekvenserne fra andre Lutra-arter, hvoraf otte kloner havde den samme nukleotidsekvens (repræsenteret ved sekvensen af L. nippon (Ehime) – c4 i Fig. 1) men den resterende (sekvensen af L. nippon (Ehime)-c5 i Fig. 1) var forskellig i to positioner (21: C-T T og 92: T-T). De elleve kloner havde cytochrom b-lignende sekvenser som beskrevet nedenfor. De resterende 10 kloner havde uventet den næsten identiske sekvens til svinecytochrom B-sekvensen, hvilket tyder på, at der opstod en alvorlig DNA-kontaminering under opbevaring af den mumielignende prøve.
vi fandt ud af, at DNA ‘ et fra en mumielignende Lutra nippon (Ehime) indeholder en cytokrom b-lignende sekvens. Overraskende svarede alle de forstærkede produkter fra en anden Lutra nippon fra Kochi (mindst 30 år gammel udstoppet prøve) til de cytokrom B-lignende sekvenser, og intet mitokondrielt cytokrom B-fragment blev forstærket. Derudover blev de cytokrom B-lignende fragmenter på 307 bp forstærket med et udbytte på 30-40% af de samlede kloner fra DNA ‘erne fremstillet af den friske lever eller hår fra de eurasiske oter L. lutra (Europa og Asahikava), men ikke fra DNA’ erne fra L lutra (Letland og Kina). Tilstedeværelsen af cytokrom B-lignende sekvens er således ikke en artefakt under en lang opbevaring af DNA. Den cytokrom b-lignende sekvens findes også i det nukleare DNA i en gnaver (Smith et al., 1992). De cytochrome b-lignende sekvenser af Lutra er kendetegnet ved tilstedeværelsen af en sletning for at forårsage en læserammeforskydning og termineringskodoner (Fig. 1), ved tilstedeværelsen af flere typer sekvenser med mindre variationer (Fig. 1) og ved den tilsyneladende hurtigere evolutionære hastighed sammenlignet med mitokondrie cytochrom B-sekvensen (se Fig. 2). Således ser den cytokrom b-lignende sekvens ikke ud til at fungere og er til stede som en pseudoform. Vi har ikke undersøgt, om den cytokrom b-lignende sekvens af Lutra er til stede i mitokondrie-DNA eller i det nukleare DNA.
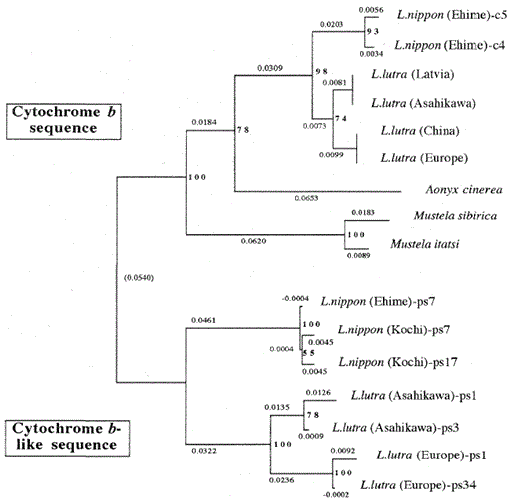
Fig. 2.
et fylogenetisk træ konstrueret ud fra sekvensjusteringen (baseret på sekvens 1-224 i Fig. 1) af mitokondrie cytochrom b og cytochrom b-lignende sekvenser med nabo-Sammenføjningsmetoden. Tallene ved hvert forgreningspunkt viser bootstrap-værdier (100 replikationer), ps repræsenterer den cytokrom b-lignende sekvens.
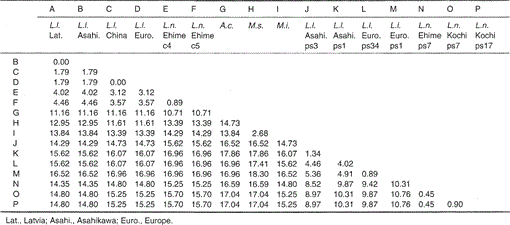
nucleotidsekvenserne af mitokondrie cytochrom b og cytochrom b-lignende gener af Lutra nippon er på linje med dem af L. lutra (Letland, Kina, Europa og Asahikava), aonyks cinerea, Mustela sibirica og Mustela itatsi, i Fig. 1. Af sekvensen 1 til 224 var Lutra nippon (C4) forskellig fra L lutra (Letland) og L. lutra (Kina) i henholdsvis 9 og 7 nukleotider, som alle var placeret i den tredje position af et kodon og identificeret som overgangsforskelle a Lutra G eller C C. Forskellen inden for Lutra lutra var 4 nukleotider. Disse nukleotidsubstitutioner forårsager ingen aminosyreudskiftning. På den anden side er der 6 nukleotider forskel i de to medfødte væsler Mustela sibirica og Mustela itatsi i denne region. Således er sekvensforskellen mellem Lutra nippon og Lutra lutra (7-9 nukleotider: 3,6% i gennemsnit) større end den (6 nukleotider) mellem de to Mustela-arter. Dette er også i overensstemmelse med tanken om, at I Mustelidae-arten kan mere end 3,5% forskel i sekvens af cytokrom B-gen svare til forskellen mellem forskellige arter (Masuda og Yoshida, 1994). Den procentvise forskel mellem nukleotidsekvenserne er vist i tabel 1.
tabel 1.
procentforskel mellem nukleotidsekvenserne (224 bp) vist i Fig. 1
figur 2 viser et fylogenetisk træ konstrueret ud fra sekvensdata opnået i denne undersøgelse med nabo-Sammenføjningsmetoden i PHYLIP-pakkeversionen 3.5 c (Felsenstein, 1993). Den samme topologi blev rekonstrueret med Fitch og Margoliash-metoden (Felsenstein, 1993). Sekvenserne blev opdelt i to hovedklynger, den ene er for mitokondrie cytokrom B-gener og den anden for cytokrom b-lignende sekvenser. Som nævnt ovenfor blev de cytochrome B-lignende sekvenser forstærket fra DNAs af Lutra nippon (2 individer, fra Kochi og Ehime), L. lutra (Europa) og L. lutra (Asahikava). Forgreningsmønsteret for de cytokrom B-lignende sekvenser af de to Lutra-arter var det samme som i mitokondrielle cytokrom B-gener, men den evolutionære hastighed for de cytokrom b-lignende sekvenser var mindst to gange hurtigere, i overensstemmelse med de generelle egenskaber ved pseudogener. Træet viser også tydeligt, at der er en stor genetisk forskel mellem Lutra nippon og Lutra lutra, og dette resultat er i overensstemmelse med den nylige morfologiske undersøgelse af, at den japanske flododder ikke er en underart af den eurasiske Odder Lutra lutra, men en særskilt art, Lutra nippon (Imaisumi og Yoshiyuki, 1989). Imidlertid, for at få et afgørende bevis for det fylogenetiske forhold mellem de to arter, længere DNA-sekvens af Lutra nippon skal bestemmes. Dette ville ske, når den friske prøve af den japanske flododder blev opnået.