소개
몽골어로 치게,치고어,아라그 또는 아이 라그라고도 불리는 쿠미스는 전통적인 발효 유제품의 한 유형입니다. 그것은 수세기 동안 중국의 몽골과 내몽고에서 인기있는 음식이었습니다(장과 장,2011). 이 지역의 사람들은 대축제와 희생 제물(장과 장,2011)동안 쿠미스를 소비했습니다. 쿠미스 생산의 가장 초기 기록은 한 왕조(기원전 202 년~서기 202 년)로 거슬러 올라갈 수 있습니다. 이 제품은 위안 왕조(서기 1271-서기 1368)(장 외)동안 널리 인기를 얻었습니다. 2010 년). 오늘날 쿠미스는 몽골과 내몽고 지역 주민들에게 흔한 식품이지만,이 지역 중 소수만이 산업 규모로 제조됩니다. 쿠미스는 높은 함량의 필수 아미노산과 비타민을 포함한 풍부한 영양소를 제공 할뿐만 아니라 광범위한 의학적 상태를 완화시키는 것으로 여겨지며 수술 후 치료에 유익합니다(야기 엘 스키,1877;톰슨,1879).
전통적으로 쿠미스는 일반적으로 나무 통,동물의 피부로 만든 용기 또는 항아리에서 생산됩니다. 발효는 스타터 문화(장과 장,2011)의 역할 오래된 쿠미스와 용기에 여과 암말 우유를 첨가 한 후 주변 온도에서 자연적으로 발생합니다. 쿠미스는 생명 공학 잠재력의 새로운 박테리아의 좋은 소스입니다(장 외. 2010;팬 외., 2011). 따라서 가능한 한 많은 발효 관련 쿠미스 박테리아를 탐구하고 보존하는 것이 매우 중요합니다. 지난 수십 년 동안,연구의 수는 쿠미스 세균 지역 사회를 조사하기 위해 수행되었다(우 외. 2009;하오 외.,2010),주로 문화-,분자 생물학-및 파이로 시퀀스 기반 방법에 의해 연구(일 외., 2010). 이러한 다양한 접근법 중 파이로 시퀀스 기반 방법은 표현형 특성 및 개별 미생물의 배양 가능성 문제로부터 독립적 인 쿠미스의 가장 포괄적 인 미생물 군 프로파일을 제공했습니다. 그러나 쿠미스 박테리아에 의해 코딩 된 기능성 유전자의 스펙트럼과 발효 능력은 특히 희귀 한 미생물 집단에 대해 특성화되지 않았습니다.
본 연구는 단일 세포 유전체학 기술을 사용하여 중국 몽골과 내몽골에서 수집 한 10 개의 쿠미스 샘플의 박테리아 메타 게놈을 분석했습니다. 현재 작업 낙농 제품에 세균 다양성 조사에 최첨단 기술을 적용 했습니다. 우리의 연구는 단일 셀 메타 유전체학 접근 방식을 적용 하 여 낮은 풍부한 분류군을 발견의 타당성을 입증 했다. 고무적인 결과는 전통적인 연구 분야의 문제를 해결하는 새로운 접근법의 개발 및 적용을 촉진 할 것입니다.1741>
재료 및 방법
샘플의 제조
메타게노믹스 연구를 위해 몽골과 중국 내몽골에서 총 10 개의 쿠미스 샘플을 수집하였다. 샘플 무 균 적으로 수집 하 고 드라이 아이스에서 수송 했다.
각 샘플의 1 밀리리터는 와드 등에서 기술된 방법에 따라 전처리되었다. (2013)일부 수정. 간단히,샘플을 3-5 분 동안 얼음 욕조에서 해동시켰다. 샘플이 녹은 후,그들은 불순물 및 진핵 세포 덩어리를 제거하기 위해 저속 원심 분리를 받았다. 그런 다음 원핵 세포를 우유 혈청으로부터 원심 분리에 의해 13,000 에서 15 분 동안 펠렛 화 하였다. 상기 펠렛을 1%트리톤 엑스-100 으로 2 밀리리터의 인산염 완충 식염수에 재 부유시키고,나머지 진핵 세포를 용해시키기 위해 2 시간 동안 배양 하였다. 그 후,박테리아 산탄에 의한 원심분리에서 13,000×g15min 와 펠릿은 다시 중단에서 500µL PBS. 마지막으로,원심 분리 단계를 한 번 더 반복하여 박테리아 세포를 세척 하였다.
구배 희석 및 다중 변위 증폭
저 풍부한 박테리아를 검출하기 위해,각 쿠미스 샘플로부터 유도 된 박테리아 현탁액을 후속 증폭 반응을 위해 직렬로 희석시켰다. 각 샘플의 셀 번호는 셀 카운팅 챔버(중국 상하이 치우징)를 사용하여 현미경(일본 도쿄 니콘)으로 대략 추정되었습니다. 희석 단계는 각 박테리아 현탁액의 세포 수가 약 100 에 도달 할 때까지 계속되었다. 묽게 한 세포의 다수 진지변환 증폭은 제조자의 지시에 따라 단세포 장비(미국 메릴랜드 주 저먼타운)를 사용하여 실행되었습니다.
도서관 구축 및 시퀀싱
증폭 된 유전자 무작위로 전단 하 고 약 500 혈압의 단편을 선택 했다. 도서관 구축 후,퍼키넬머 랩칩,지엑스 터치 및 스텝원플러스트 실시간 회로 시스템을 사용하여 도서관 품질 검사를 실시하였다. 마지막으로,125 혈압 쌍 엔드 읽기는 제조업체의 지침에 따라 일루미나 하이세크 2500 플랫폼에서 시퀀싱되었다.
데이터 분석
시퀀스 품질 검사 및 필터링
시퀀서에 의해 생성된 원시 읽기에는 라이브러리 생성 중 어댑터 오염의 인공 읽기가 포함될 수 있습니다. (1)어댑터 오염으로 인한 읽기 제거; (2)평균점수가 30 분기의 최고점수보다 낮은 판독값의 제거,(3)”엔”이 상당히 초과된 판독값의 제거(2)판독값의 5%). 다운스트림 분석은 깨끗한 데이터를 기반으로 했습니다. 또한 30 분기와 30 분기를 기준으로 통계적 기준 품질을 계산했습니다.
숙주 게놈에 대한 정렬을 수행하여 숙주 유래 오염물 서열을 제거하였다. 모든 호스트 유래 읽기는 박테리아(또는 바이러스)게놈 참조 서열과 더 비교하기 전에 폐기되었습니다. 보다 정확한 결과를 얻기 위해 버로우–휠러 얼라이너(버전 0.97)모델이 선형(리 및 더빈,2009)에 사용되었습니다.
분류학적 할당 및 다양성
웹 소프트웨어 메타플란은 속 및 종 수준에 대한 분류학적 할당에 사용되었다(세가타 외., 2012). 샘플 내 및 샘플 간의 종의 다양성을 비교 하기 위해 우리는 연구 관련 패키지에 의해 알파 및 베타 다양성을 분석 했다.
읽기 어셈블리,유전자 예측 및 주석
보다 포괄적인 정보를 얻으려면,우리는 게놈(콘티그)에 전단된 단편을 조립. 그러나,메타 게놈 샘플에서 일반적인 여러 종의 존재로 인해,우리는 스페이드(버전 3.6.2),사내 스크립트 및 메타 게놈 데이터베이스(제르 비노와 버니,2008;누크 등)를 통합하여 일반적으로 단일 종 분석에 사용되는 생물 정보학 게놈 조립 방법을 개선., 2013).
메타젠마크 소프트웨어는 조립된 콘티그(노구치 외)의 유전자 예측에 사용되었다., 2006). 그 결과,유전자는 유전자뿐만 아니라 유전자뿐만 아니라 유전자에도 영향을 미칩니다., 2012). 유전자의 상대적 풍부는 동일한 절차를 사용 하 여 유전자 카탈로그에 고품질 시퀀싱 읽기를 정렬 하 여 결정 했다. 다운스트림 불일치 분석 유전자 상대적 풍부도에 근거 했다. 유전자 주석 여러 공용 데이터베이스에 대 한 고품질 시퀀스를 정렬 하 여 수행 되었다(즉 비 중복 데이터베이스,번호,단백질,톱니의 직교 그룹의 클러스터; 유전자와 게놈의 교토 백과 사전,케그)폭발을 사용하여(알츠철 등., 1997). 도메인 검색은 인터포스캔(멀더 및 애프바일러,2008)을 사용하여 수행되었습니다.
뉴클레오티드 서열 가입 번호
본 연구에서보고 된 서열 데이터는 서열 데이터베이스에 기탁되었다.083102).
결과
실험 설계 및 시퀀싱
쿠미스에서 낮은 풍부한 박테리아를 검출하기 위해 단일 세포 증폭 기술을 사용하여 샘플의 메타 게놈을 분석했습니다. 3 개의 박테리아 현탁액,각각 약 100 개의 세포는 직렬 희석에 의해 독립적 인 쿠미스 샘플에서 파생되었습니다. 총 30 개의 희석 된 현탁액을 분석 하였다. 각각의 희석 된 샘플에는 다른 샘플 코드,즉 샘플 식별 번호 다음에 1,2 또는 3 이 주어져 3 개의 개별 희석을 나타냅니다. 일부 희귀 종 희석 중 하나에 존재 하는 전제를 바탕으로,세포의 여러 변위 증폭 실시 했다;그리고 모든 쿠미 스 박테리아 서 스 펜 션에 대 한 약 5 기가 바이트 데이터를 생성 했다.
총 1,040,323,864 개의 원시 읽기가 10 개의 쿠미스 샘플(총 30 개의 박테리아 현탁액)에서 생성되었습니다. 각 100 셀 서스펜션에 대한 평균 읽기 수는 34,677,462 입니다. 정규화되지 않은 서열을 트리밍하고 필터링 한 후 모든 샘플에 대해 1,018,381,702 개의 깨끗한 읽기를 얻었습니다. 섀넌 지수,심슨 지수,차오 1 지수 및 관찰 된 종의 수(그림 1-4)의 값은 대부분의 쿠미스 샘플이 박테리아 생물 다양성이 높다는 것을 보여주었습니다. 섀넌-위너 다이버 시티 곡선은 시퀀스 깊이가 모든 샘플에 적합하다는 것을 보여주었습니다(그림 1).
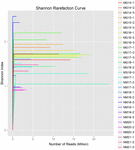
그림 1. 쿠미스 샘플의 미생물 다양성을 추정 섀넌 희박화 곡선.
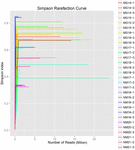
그림 2. 쿠미스 샘플의 미생물 다양성을 추정 심슨 희박 곡선.
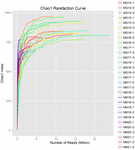
그림 3. 쿠미스 샘플의 미생물 다양성을 추정하는 차오 1 희박 곡선.
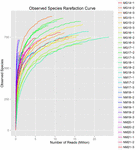
그림 4. 쿠미스 샘플의 미생물 다양성을 추정 관찰 종 희박 지수.
분류학 주석
높은-품질 시퀀스 샘플 세균 커뮤니티의 심층 분석을 사용 하 여 다른 분류학 수준에 할당 되었습니다. 쿠미스 생물 다양성에 대한 일부 발표 된 연구를 참조하여(우 외. 2009;하오 외.,2010;일 외.,2010),우리는 알려진 것과 이전에보고되지 않은 쿠미 스 관련 박테리아를 각각 공통 및 희귀 분류군으로 분류했습니다.
고품질 시퀀스는 13 개의 다른 속을 나타냅니다(그림 5). 그 중 3 개는 유산균(엘.),락토 코커스 및 연쇄상 구균을 포함하여 평균 상대 풍부도가 1%이상이었습니다. 특히,락토 바실러스와 락토 코커스는 쿠미스에서 발견 된 가장 풍부한 두 속이었다. 샘플에서 락토 바실러스의 비율은 52.72 에서 99.96%까지 다양했다. 이 속의 두 구성원,엘.헬 베티 쿠스 과 엘.케 피라 노 파시 엔스,대부분의 쿠미스 샘플 중에서 지배적이었습니다(그림 6). 종 엘. 부흐네리는 주로 몽골 샘플에 존재했고,락토코커스 락티스는 샘플링 영역에 관계없이 대부분의 샘플에서 검출되었다.
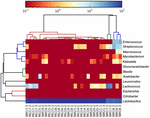
그림 5. 속 수준에서 쿠미스 샘플에서 검출 된 박테리아의 상대적 풍부를 보여주는 히트 맵.
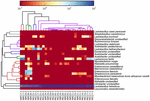
그림 6. 종 수준에서 쿠미스 샘플에서 검출 된 박테리아의 상대적 풍부를 보여주는 히트 맵.
메타 게놈 어셈블리,유전자 예측 및 기능적 주석
판독값의 조립은 614,392,623 혈압의 조립 길이를 초래했다. 어셈블리의 값은 5,596 에서 35,200 사이였습니다. 쿠미스 샘플에서 예측된 유전자의 수는 10,347 에서 34,547 사이였으며,평균 길이는 647.82 에서 985.33 사이였다. 경험적 유전자 기능 분석 현재 연구의 범위를 넘어 했다,비록 우리가 주로 시퀀스 상 동성에 따라 유전자 기능을 예측 하는 코그와 케그 데이터베이스를 사용 하 여 쿠미 스 박테리아 미생물 군집을 주석 달았다.
주석 출력에서 총 545 일치 유 당 이용 유전자(탄수화물과 신진 대사의 톱니 범주,지),높은 상 동성을 보였고 그들 중 일부는 동일한 콘티 그(표 1)내에 위치 했다.
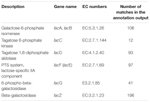
표 1. 쿠미스 박테리아 메타 게놈에 주석이 달린 유당 대사 관련 유전자.
일부 다른 서열은 카제인 분해 단백질 분해 효소에 해당하는 서열을 포함하여 아미노산 수송 및 대사의 톱니 범주 내의 상 유전자를 코딩 할 수 있으며,4-18 잔기의 올리고 펩타이드 및 아미노 펩 티다 제(예:류실 아미노 펩 티다 제,펩 티딜-디펩 티다 제 에이,아미노 펩 티다 제 엔,프롤린 이미 노 펩 티다 제 치료 및 치료 일부 서열은 아르기닌,아스파 테이트,메티오닌 및 이소 루이 신에 대한 아미노 트랜스퍼 라제 특이 적과 높은 유사성을 공유했지만,하나의 중요한 일치 만이 확인되었으며,이는 티로신 및 페닐알라닌에 대한 아미노 트랜스퍼 라제 특이 적입니다. 마지막으로,에스-리보실호모시스테인 리아제,아르기니노숙시네이트 리아제,아스파테이트 암모니아-리아제,시스타티오닌 감마-리아제,히스티딘 암모니아-리아제,및 오-아세틸호모세린(티올)-리아제를 포함하는 다수의 서열이 아미노산 리아제에 높은 상동성을 공유하였다.
토론
쿠미스는 몽골과 중국 내몽골에서 인기있는 전통 발효 유제품입니다. 쿠미스의 박테리아 다양성을 조사하기 위해 많은 연구가 수행되었지만 쿠미스 미생물의 유전 적 용량에 관한 정보는 거의 얻지 못했습니다. 여기,우리는 특히 낮은 풍부한 세균 인구에 초점을 맞추고 쿠미 스 박테리아 메타 게놈을 분석 하는 단일 셀 증폭 기법을 적용.
전형적인 메타 게노믹스 접근법은 이전에 쿠미스 미생물을 설명하기 위해 적용되었습니다. 그러나 깊은 서열을 달성하는 데 드는 높은 비용으로 인해 샘플의 희귀 미생물 군집은 종종 부적절하게 덮여 있습니다. 따라서 소수 박테리아의 계통 발생 및 기능성 메타 게놈은 모두 제한적입니다. 우리의 단일 셀 증폭 방법 100 셀 현 탁 액 샘플의 직렬 희석을 포함 했다. 희석 된 쿠미스 샘플에 존재하는 세포의 수가 적기 때문에 제한된 양의 유전자 만 추출됩니다. 증폭 단계는 분석될 유전자 물질의 양을 증가시켰고,따라서 분 유전자 양을 포함하는 샘플의 메타게놈 분석을 촉진시켰다. 이 방법의 한 가지 단점은 각 분류군의 시퀀스 과정에서 동등 하 게 증폭 될 것 이라고 보장에 어려움; 따라서 여기에서 얻은 결과는 시퀀스의 상대적 풍부도 만 반영 할 수 있지만 확인 된 분류군 또는 기능 유전자의 절대 양은 반영 할 수 없습니다. 아직,이 우리의 분석에 거의 영향을 미칠 것 이다 현재 연구 다른 출판된 작품 희귀 세균 인구를 초점에서 다릅니다. 이 작업에 의해 생성 된 데이터는 과소 대표 된 인구에게 보완적인 정보를 제공합니다. 우리는 현재의 접근 방식은 생태 환경의 다른 유형에 대한 박테리아 다양성의 미래 분석에 적합하다고 생각합니다.
쿠미스 박테리아 미생물은 주로 유산균(실험실)과 일부 아세트산 박테리아(장 및 장,2011)로 구성됩니다. 예상대로,우리의 데이터 세트는 대부분 실험실 및 아세트산 박테리아를 나타내는 시퀀스를 포함. 락토 바실러스 헬베 티 쿠스에 해당하는 서열은 모든 샘플에서 지배적이었습니다. 또한,종을 나타내는 서열,엘.케 피라 노 파시 엔스. 엘 부흐네리 엘.케피라노파시엔스. 장구균(이자형)카셀리프라부스. 페칼리스 전자. 류코노스토크 장간막종 락토 코커스 락 티스,및 아세토 박터 저온 살균,또한 발견되었다. 다른 분류군의 시퀀스의 식별 박테리아의 생존 능력을 보여 충분 하지 않을 수 있습니다.,그럼에도 불구 하 고 발효 과정의 어떤 시점에서 세균 커뮤니티를 반영. 미야모토 외. (2010)는 마지막 거치한 제품에 있는 세균성 보급이 그들의 산성 긴장 포용력과 관련있다는 것을 건의합니다. 일반적으로,유산균은 우리의 데이터 세트에 존재하는 유산균 서열의 높은 상대적 풍부를 설명 할 수있는 유산균보다 높은 산 내성을 갖는다. 다른 한편으로는,리터의 빈번한 발생. 쿠미스의 헬베티쿠스는 엘.헬베티쿠스 서열의 우위의 관찰과 일치.오타 키엔 시스,이는 쿠미스 또는 다른 유제품 틈새에서보고 된 적이없는 희귀 종입니다. 이 종은 먼저 일본의 전통 피클 인 선키 생산에 사용 된 비 염장 절임 용액으로부터 설명되고 분리되었습니다(와타나베 외., 2009). 그것은 레카 유전자를 기반으로 한 증폭 된 단편 길이 다형성 프로파일 링에 의해 발견되었습니다. 그 이후로,이 종은 다른 식품 관련 틈새 시장과 관련이있는 것으로보고되지 않았습니다. 따라서,이 박테리아는 피클의 자생 식물 군에 속할 가능성이 높습니다. 그러나,우리는 그것이 채택된 검출 방법의 감도 때문에 단순히 찾아내지 않았다 가능성을 배제할 수 없습니다. 락토바실러스 오타키엔시스는 디-류신,디-알로-이소류신 및 디-발린(도이 외)과 같은 디-분지 사슬 아미노산을 생산할 수 있다., 2013). 그것은 특정 발효 식품의 생산 특성 향상에 사용될 가능성이있다(카토 외., 2011).
우리의 연구는 또한 쿠미스 관련 박테리아로보고 된 적이없는 종의 마케도니쿠스를 나타내는 서열을 확인했다. 대신,그것은 그리스 양과 염소 치즈에 존재하는 스타터 문화입니다(게오 갈라 키 외., 2000). 이 종의 일부 구성원은 엑소 폴리 사카 라이드,박테리오신(빈센트 외)을 생산할 수 있습니다. 2001;아나스타시우 외. 2015),및 감마-아미노 부티르산(프란치오시 외., 2015). 이 종은 발효 식품에서 자주 분리되지만,원래 틈새 시장 에스., 2016). 아니 최근까지,파파디미트리우 외. 또한,락토 코커스 락 티스로부터 획득 된 플라스미드를 확인 하였다. 플라스미드는 낙농 제품 환경 내에서 조상의 유전 적 교환 사건을 통해 옮겨 졌을 가능성이 있으며,이는 낙농 기원을 암시합니다., 2015). 호모 필 루스. 이 병리의 주요 원인 중 하나는 병원균,병원균,병원균,병원균,병원균,병원균,병균,병균,병균,병균,병균,병균,병균,병균,병균,병균,병균,병균,병균 및 병균이다.
이 박테리아는 일반적으로 장내 환경에서 발견되기 때문에 루미 노 코커스 토르크 종을 나타내는 서열의 식별은 예상치 못한 것이었다. 그것은 분비 글리코시다 제의 구성 생산에 의해 점액 올리고당을 저하시킬 수있는 정상적인 인간의 장 미생물입니다(호스킨스 외., 1985). 최근 임상 증거는이 종의 배설물 풍부 자폐 스펙트럼 장애;와 어린이에서 변경 됩니다 보여줍니다 그러나,장애에서 그것의 역할은 불분명(왕 외., 2013).
놀랍게도,일부 서열은 잠재적인 병원체를 나타냈다. 예를 들어,클렙시 엘라 폐렴은 인간 및 동물 내장의 약 40%에 서식하는 기회 주의적 인간 병원체입니다. 이전에 오릭스 바실러스라고 불리는 마이코박테리움 오리기스는 인간 결핵을 일으킬 수 있는 마이코박테리움 결핵 복합체의 일원이다(도슨 외., 2012). 이 두 종은 원시 우유에서보고되었지만 쿠미스는보고되지 않았습니다. 따라서 이들의 존재는 특히 비 무균 또는 저 무균 조작 조건 하에서 기존의 쿠미스 생산 중 오염에 기인 할 수 있습니다.
박테리아 대사는 쿠미스 특성과 품질을 형성하는 데 중요한 역할을합니다. 지방 분해 및 단백질 분해와 같은 미생물 기반 공정은 쿠미스 방향족 및 향료 화합물을 합성하는 데 필요합니다., 2016). 일관되게,박테리아 메타 게놈은 잠재적으로 유당 분해 및 단백질 분해 시스템을 코드화하는 서열을 포함했습니다. 상대적으로 간단한 유당 이화 작용 통로와는 다른,실험실 단백질 분해 체계는 효소의 다양한 배열로 위로 만듭니다(첸 외., 2014). 다른 검출 된 쿠미스 실험실과 비교하여,우성 종 엘.헬베티쿠스는 높은 단백질 분해 활성을 특징으로한다(장 외., 2015). 대부분의 실험실은 우유 카제인 가수 분해를 시작하는 단 하나의 세포 외피 단백질 분해 효소를 가지고있는 반면,엘.헬 베티 쿠스는 이들 효소 중 적어도 두 가지를 포함합니다., 2011). 따라서,엘.헬 베티 쿠스 및 단백질 분해 관련 유전자의 강한 단백질 분해 종에 해당하는 서열의 높은 비율은 쿠미스에서 펩티드 및 유리 아미노산의 비교적 높은 함량과 연결될 수있다.
산업 규모 쿠미스 생산의 한 가지 어려움은 쿠미스가 전통적으로 자연 발효에 의해 만들어 졌기 때문에 풍미 인식의 제어입니다. 따라서,특히 천연 오염 물질의 존재 하에서,주요 풍미 성분의 풍미 및 프로파일을 정의하는 것은 어려웠다. 주요 맛 성분의 생산은 분지 사슬 아미노산,메티오닌 및 방향족 아미노산과 같은 아미노산의 발효 및 효소 분해 결과입니다(아르도,2006). 향미 성분의 예로는 알데히드,유기산 및 에스테르가 있으며,이는 트랜스 아미나 제(에서)-경로(헬링크 외)에 의해 형성된다., 2004). 이 경로는 아미노산이 해당 아미노산으로 전환되는 것을 촉매하는 트랜스 아미나 제에 의해 시작됩니다., 2004). 우리의 데이터 세트의 일부 서열은 추정 분지 사슬 및 방향족 아미노산 아미노 전이 효소에 해당합니다. 특히,우리는 쿠미스 관련 종의 콘티그에서 추정 방향족 아미노산 아미노 전이 효소 나는 발견,에스. 우리의 현재 연구는 실리코의 마이크로 바이 옴에만 주석을 달았 기 때문에,우리의 데이터는 이러한 확인 된 유전자 서열이 실제로 앞서 언급 한 쿠미스 향미 화합물을 생산하는 데 실제로 기능적이라는 것을 보여주기에 충분하지 않습니다. 이러한 유전자의 존재는 그럼에도 불구 하 고 그들은 같은 발효 활동;에 대 한 몇 가지 가능한 후보 제안 아직,추가 실험 작업 그들의 정확한 기능적 역할을 명료 하 게 하는 데 필요한 것 이다.
또한,우리는 아미노산 리아제 및 트레오닌 알 돌라 제를 포함한 다른 아미노산 전환 경로에 해당하는 서열을 발견했다. 이전의 효소는 메티오닌을 메탄 티올로 전환시키는 촉매 작용을한다. 디메틸 디설파이드 및 디메틸 트리설파이드 형성(페르난데스 외. 트레오닌이 글리신 및 아세트 알데히드로 전환되는 것을 촉매하는 반면,후자는 트레오닌을 글리신 및 아세트 알데히드로 전환시키는 것을 촉매한다., 2000). 마찬가지로,단지 쿠미스 마이크로 바이 옴 내에서 이러한 서열을 찾는 것은 실제 생물학적 역할에서 확실한 증거로 작용할 수 없다;미래의 실험적 확인이 필요할 것이다.
결론
본 연구는 중국의 몽골과 내몽골에서 수집 한 쿠미스 샘플의 박테리아 마이크로 바이 옴을 분석하기 위해 변형 된 메타 게노믹스 방법을 사용했다. 우리는 쿠미스;희귀 종의 계통 및 기능 메타 게놈 특징 그리고 우리의 데이터 집합 생명 공학 관심과 잠재력의 특성을 반영 한다. 우리의 연구는 처음으로 미생물 오염 물질 뿐만 아니라 쿠미스 박테리아 미생물 검출에 단일 셀 증폭 기술을 통합의 타당성을 입증 했다. 본원에서 개발 된 기술은 소수 미생물 인구에 초점을 맞춘 발효 과정을 따라 쿠미스 마이크로 바이 옴의 변화를 모니터링하기 위해 향후 연구에서 사용될 수 있습니다. 또한,쿠미스 마이크로 바이 옴의 기능 및 대사 능력을 확인하기 위해 전사 체학,대사 체학과 같은 다른 오믹스 기술을 현재의 메타 게노믹스 분석에 결합 할 수 있습니다.
기술적으로,이 작업의 다음 단계는 현재 방법을 최적화하는 것입니다. 예를 들어,유전자 증폭 전에 샘플 희석을 증가시킴으로써 희귀하고 새로운 박테리아를 발견 할 수있는 기회가 향상 될 수 있습니다. 한편,태평양 단일 분자 생명 과학,실시간 시퀀싱 기술로 긴 읽기를 생성 할 수있는 대체 시퀀싱 기술은 게놈 조립 공정을 개선하기 위해 사용될 수있다.
저자 공헌
이 연구를 설계했다. 원고를 작성했습니다. 1990 년대 초반부터 1990 년대 초반까지. 데이터 분석. 모든 저자는 원고를 검토했습니다.
이해 상충 성명
저자는 연구가 잠재적 인 이해 상충으로 해석 될 수있는 상업적 또는 재정적 관계가없는 상태에서 수행되었다고 선언합니다.
승인
이 연구는 중국 국립 자연 과학 재단(보조금 제 31571815 호)의 지원을 받았다.
보충 자료
은 보충 자료를 위해 이 문서에서 찾을 수 있습니다 온라인: https://www.frontiersin.org/article/10.3389/fmicb.2017.00165/full#supplementary-material
알트츌,S.F.,Madden,T.L.,Schaffer,A.,장,J.,장,Z.,Miller,W.,외. (1997). 갭 블라스트 및 사이오닉 블라스트:차세대 단백질 데이터베이스 검색 프로그램. 핵산 레.25,3389-3402. 2017 년 10 월 10 일(토)~2018 년 10 월 10 일(일)~2018 년 10 월 10 일(일)~2018 년 10 월 15 일(일)~2018 년 10 월 15 일(일)~2018 년 10 월 15 일(일)
이 경우,상기 제 1 항은 제 2 항 및 제 2 항과 제 2 항과 제 2 항을 포함한다. (2015). 삼투압 스트레스 내성 표현형을 향한 연쇄상 구균 마케도니쿠스의 조작 된 균주는 고 삼투압 조건 하에서 박테리오신 마케도신을 생산하는 능력을 유지합니다. 바이오 테크놀로지. 212, 125–133. 도이:10.1016/제이.2015.08.018
출판 초록|크로스레프 전체 텍스트|구글 학자
아르도,과.(2006). 아미노산 이화 작용에 의한 풍미 형성. 바이오 테크놀입니다. 교재 24,238-242. 도이:10.1016/제이.2005.11.005
출판 초록|크로스레프 전체 텍스트|구글 학자
2018 년 12 월 1 일,2018 년 12 월 1 일,2018 년 12 월 1 일,2018 년 12 월 1 일,2018 년 12 월 1 일,2018 년 12 월 1 일 (2014). 탈지유에서 성장하는 동안 유산균 헬베티쿠스의 프로테옴 분석. 제.유제품 과학. 97, 7413–7425. 10.3168.2014-8520
출판 초록|크로스레프 전체 텍스트|구글 학자
2012 년 12 월 15 일,2013 년 12 월 15 일,2013 년 12 월 15 일,2013 년 12 월 15 일,2013 년 12 월 15 일,2013 년 12 월 15 일,2013 년 12 월 15 일,2013 년 12 월 15 일. 결핵 환자에서 뉴질랜드의 젖소로의 마이코 박테리아 오리 기스(엠.결핵 복합 종)의 전염. 제이 클린 마이크로바이올 50, 3136–3138. 도이:10.1128/제이씨엠.01652-12
출판 초록|크로스레프 전체 텍스트|구글 학자
본 발명의 실시예는,도이,케이,모리,케이,무타구치,와이이,타시로,케이,후지노,와이이,오모리,티 등이 있다. (2013). 게놈 서열 초안 디-분지 사슬 아미노산 생산자 유산균 오타 키엔 시스 15040 티,전통적인 일본 피클에서 분리. 게놈 아나운서. 2015 년 11 월 13 일-2015 년 11 월 13 일 도이:10.1128/게노메아.00546-13
출판 초록|크로스 레프 전체 텍스트|구글 학자
이 경우,상기 제 1 항은 제 2 항 및 제 2 항과 제 2 항 및 제 2 항과 제 2 항 및 제 2 항과 제 2 항 및 제 2 항과 제 2 항 및 제 2 항과 제 2 항 및 제 2 항과 제 2 항 및 제 2 항과 제 2 항 및 제 2 항과 제 2 항 및 제 2 항과 제 2 항 및 제 2 항과 제 2 항 및 제 2 항과 제 2 항 (2000). 시스타 티오 닌 베타-리아제를 코딩하는 락토 코커스 락 티스의 메트 유전자의 분자 및 기능 분석. 신청 환경. 마이크로바이올 66, 42–48. 10.1128.66.1.42-48.2000
PubMed 추상|CrossRef 전체 텍스트|Google 학술검색
Franciosi,E.,카라파,I.,Nardin,T.,Schiavon,S.,Poznanski,E.,Cavazza,A.et al. (2015). 전통적인 알파인 생 우유 치즈에서 분리 된 유산균에 의한 생물 다양성 및 감마-아미노 부티르산 생산. 바이오메드 2015, 625740. 도이: 10.1155/2015/625740
째챈쨩처쨀짼쨉쨉짹쨀째철쩔첩쨩
2012 년 12 월 15 일-2012 년 12 월 15 일. 차세대 시퀀싱 데이터를 클러스터링하기 위해 가속화되었습니다. Bioinformatics 28, 3150–3152. doi: 10.1093/bioinformatics/bts565
PubMed Abstract | CrossRef Full Text | Google Scholar
Georgalaki, M. D., Sarantinopoulos, P., Ferreira, E. S., De Vuyst, L., Kalantzopoulos, G., and Tsakalidou, E. (2000). Biochemical properties of Streptococcus macedonicus strains isolated from Greek Kasseri cheese. J. Appl. Microbiol. 88, 817–825. doi: 10.1046/j.1365-2672.2000.01055.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Gesudu, Q. M., Zheng, Y., Xi, X. X., Hou, Q. C., Xie, H. Y.,황,더블류 큐,등. (2016). 단일 분자 실시간 시퀀싱을 사용 하 여 내몽골에서 전통적인 쿠미스에서 세균 인구 구조 및 역학 조사. 제.유제품 과학. 99, 7852–7863. 10.3168.2016-11167
출판 초록|크로스레프 전체 텍스트|구글 학자
이 경우,본 발명의 실시예에 따른 실시예는 다음과 같다. (2016). 피도 페코리노 시칠리아노 치즈 생산을 위해 선택된 자생 유산균의 대규모 공장 규모 적용. 식품 마이크로 바이올. 59, 66–75. 10.1016/2016.05.011
PubMed 추상|CrossRef 전체 텍스트|Google 학술검색
오,Y.,조,L.,장,H.,자이,Z.,황,Y.,Liu,X.,et al. (2010). 구배 겔 전기 영동 및 종 특이 적 중합 효소 연쇄 반응을 변성시킴으로써 쿠미스의 박테리아 생물 다양성의 식별. 제.유제품 과학. 93, 1926–1933. 10.3168.2009-2822
출판 초록|크로스 레프 전체 텍스트|구글 학자
2010 년 11 월 15 일,2010 년 11 월 15 일,2010 년 11 월 15 일,2010 년 11 월 15 일,2010 년 11 월 15 일. 아미노산에서 방향 화합물을 생성하는 호 열성 유산균의 능력. 신청 환경. 마이크로바이올 70, 3855–3861. 10.1128.70.7.3855-3861.2004
PubMed 추상|CrossRef 전체 텍스트|Google 학술검색
Hoskins,L.C.,Agustines,M.,맥키,W.B.,Boulding,E.T.,Kriaris,M.,and Niedermeyer,G.(1985). 인간 결장 생태계의 점액 분해. 점액 당 단백질에서 혈액형 항원 및 올리고당을 분해하는 분변 균주의 분리 및 특성. 제이 클린 투자. 75, 944–953. 도이:10.1172/111795
퍼블릭 초록/크로스레프 전체 텍스트/구글 학자
2010 년 11 월 15 일,2010 년 11 월 15 일,2010 년 11 월 15 일. 시스타 티오 닌 리아제를 코딩하는 두 개의 락토 바실러스 카 세이 유전자의 복제 및 특성화. 신청 환경. 마이크로바이올 74, 99–106. 10.1128.00745-07
출판 초록|크로스레프 전체 텍스트|구글 학자
1877. 메스꺼움,구토 및 위장에 다른 음식을 보유 할 수없는 치료에 대한 쿠미스의 가치. 브롬 메드. 2,919-921. 도이:10.1136/비엠제이.2.887.919
출판 초록|크로스 레프 전체 텍스트|구글 학자
2011 년 12 월 15 일,2011 년 12 월 15 일,2011 년 12 월 15 일,2011 년 12 월 15 일,2011 년 12 월 15 일. 레드 와인과 화이트 와인의 발효 중 아미노산 농도와 미생물 군집 구조의 변화. 제이 바이오시. 바이오엥 111, 104–108. 도이:10.1016/제이.2010.08.019
출판 초록/크로스레프 전체 텍스트/구글 학자
2009. 버로우-휠러 변환으로 빠르고 정확한 짧은 읽기 정렬. 생물 정보학 25,1754-1760. 2018 년 10 월 10 일(토)~2018 년 10 월 10 일(일)~2018 년 10 월 10 일(일)~2018 년 10 월 10 일(일)~2018 년 10 월 10 일(일)
2006. 시디 히트:클러스터링 및 단백질 또는 뉴클레오티드 서열의 큰 세트를 비교하기위한 빠른 프로그램. 생물 정보학 22,1658-1659. 2018 년 11 월 1 일,서울시 강남구 테헤란로 15 길 16(역삼동),서울시 강남구 테헤란로 15(역삼동),서울시 강남구 테헤란로 15(역삼동),서울시 강남구 테헤란로 15(역삼동)
본 발명의 실시예에 따르면,본 발명의 실시예는 다음과 같다. (2010). 전통적인 몽골 발효유에서 유산균 및 효모의 변성 그라디언트 겔 전기 영동 분석. 식품 과학. 기술. 16,319-326. 도이:10.313616.319
크로스 레프 전체 텍스트/구글 학자
2010 년 11 월 15 일. 단백질 도메인 분석을위한 인터프로 데이터베이스 및 도구. 커 프로토토 생물 정보학 제 2 장,단위 2.7. 2018 년 10 월 10 일(토)~2018 년 10 월 15 일(일)
2010 년 11 월 1 일,2010 년 11 월 1 일,2010 년 11 월 1 일,2010 년 11 월 1 일. 메타진: 환경 게놈 샷건 서열에서 원핵 유전자 발견. 핵산 레.34,5623-5630. 2017 년 10 월 10 일(토)~2018 년 10 월 10 일(일)
이 경우,의사는 환자의 상태를 진단하고 진단하는 데 도움을 줄 수 있습니다. (2013). 키메라 유전체 제품에서 단일 세포 게놈과 미니 메타 게놈을 조립합니다. 컴퓨터. 바이올 20, 714–737. 도이:10.1089/센티미터.2013.0084
출판 초록|크로스레프 전체 텍스트|구글 학자
2015 년 11 월 1 일,2015 년 11 월 1 일,2015 년 11 월 1 일,2015 년 11 월 1 일 (2000). (13)를 사용하여 우유 발효 중 아세트 알데히드의 기원 씨 표지 전구체. 제이 아그릭. 식품 화학. 48, 1512–1517. 2018 년 10 월 10 일(토)~2018 년 10 월 10 일(일)
2011 년 11 월 15 일,2011 년 11 월 15 일,2011 년 11 월 15 일,2011 년 11 월 15 일,2011 년 11 월 15 일. 콜레스테롤 저하 효과가있는 잠재적 인 프로 바이오 틱 박테리아 인 쿠미스로부터 분리 된 락토 바실러스 페르멘툼의 특성화. 제이 사이언스. 음식. 농업. 91, 512–518. 10.10024214
퍼블릭 초록|크로스레프 전체 텍스트|구글 학자
파파 디미 트리 우,케이. 100,000,000,000,000,000,000,000,000,000,000,000,000,000,000,000,000,000,000,000,000,000,000,000 (2015). 이 종의 낙농 기원에 대한 198 점을 지적한다. 10:이자형 0116337. 도이:10.1371/저널.폰0116337
출판 초록|크로스 레프 전체 텍스트|구글 학자
2012 년 12 월 15 일,2013 년 12 월 15 일,2013 년 12 월 15 일,2013 년 12 월 15 일,2013 년 12 월 15 일,2013 년 12 월 15 일,2013 년 12 월 15 일,2013 년 12 월 15 일. 독특한 클래드 특이 적 마커 유전자를 사용한 메타 게놈 미생물 군집 프로파일 링. 냇 방법 9,811-814. 도이:10.1038/엔 메스.2066
출판 초록|크로스 레프 전체 텍스트|구글 학자
본 발명의 실시예는 다음과 같다. (2010). 중국에서 쿠미스로부터 분리 된 지배적 인 유산균의 식별 및 특성화. 애플 마이크로바이올 56, 257–265. 10.2323/제이감.56.257
퍼블릭 초록|크로스 레프 전체 텍스트|구글 학자
톰슨,제이(1879). 질병을 낭비하는 쿠미스의 가치. 브롬 메드. 1,270. 도이:10.1136/비엠제이.1.1466.270c
PubMed 추상|CrossRef 전체 텍스트|Google 학술검색
트 빈센트,S.J.,Faber,E.J.,Neeser,J.R.,Stingele,F.,그리고 Kamerling,J.P.(2001). 에 의해 생성 된 엑소 폴리 사카 라이드의 구조 및 특성. 글리코 생물학 11,131-139. 10.1093/글리콥/11.2.131
펍메드 초록/크로스레프 전체 텍스트/구글 학자
2013 년 12 월 15 일,2013 년 12 월 15 일,2013 년 12 월 15 일,2013 년 12 월 15 일,2013 년 12 월 15 일,2013 년 12 월 15 일,2013 년 12 월 15 일,2013 년 12 월 15 일. 수테 렐라 종의 풍부함 증가. 그리고 자폐증 스펙트럼 장애가있는 어린이의 대변에서 루미 노 코커스 토크. 몰 자폐증 4,42. 도이: 10.1186/2040-2392-4-42
째챈쨩처쨀짼쨉쨉짹쨀째철쩔첩쨩
2013 년 11 월 15 일,2013 년 11 월 15 일,2013 년 11 월 15 일,2013 년 11 월 15 일,2013 년 11 월 15 일,2013 년 11 월 15 일. 인간 우유 메타 게놈:기능적 용량 분석. 마이크로바이올 13:116. 도이: 10.1186/1471-2180-13-116
째챈쨩처쨀짼쨉쨉짹쨀째철쩔첩쨩
본 발명의 실시예에 의하면,본 발명의 실시예는 다음과 같다. (2009). 락토바실러스 키소넨시스 11 월 100%천연 유산균. 11 월 락토바실러스 라피 11 월 그 이유는 무엇입니까? 11 월,전통적인 일본 피클 인 선키에서 분리 된 이종 발효 종. 지능. 제이 시스트. 에볼 마이크로바이올 59(태평양 표준시 4),754-760. 도이:10.1099/아이즈.0.004689-0
PubMed 추상|CrossRef 전체 텍스트|Google 학술검색
Wu,R.,왕,L.,Wang,J.,Li,H.,Menghe,B.,Wu,J.,et al. (2009). 내몽고 쿠미스에서 유산균의 분리 및 예비 프로 바이오 틱 선택. 기본 마이크로 바이올. 49, 318–326. 도이:10.1002/작업.200800047
출판 초록|크로스레프 전체 텍스트|구글 학자
2010 년 11 월 1 일. 벨벳:드 브루 그래프를 사용하여 드 노보 짧은 읽기 어셈블리 알고리즘. 18,821-829. 도이:10.1101/할머니.074492.107
출판 초록|크로스 레프 전체 텍스트|구글 학자
2010 년 12 월 15 일,2010 년 12 월 15 일,2010 년 12 월 15 일,2010 년 12 월 15 일,2010 년 12 월 15 일 쿠미스로부터 분리된 락토바실러스 카세이 균주에서의 이올 클러스터의 스크리닝 및 서열 분석. 폴리아 마이크로바이올. (프라하)55,603-606. 도이:10.1007/s12223-010-0097-3
PubMed 추상|CrossRef 전체 텍스트|Google 학술검색
장,W.,Yu,D.,Sun,Z.,Wu,R.,Chen,X,Chen,W.,et al. (2010 비). 중국 내몽골의 전통적인 수제 쿠미스에서 분리 된 새로운 프로 바이오 틱 균주 인 락토 바실러스 카세이 장의 완전한 게놈 서열. 제.박테리올. 192, 5268–5269. 도이:10.1128/제이비.00802-10
펍메드 초록|크로스레프 전체 텍스트|구글 학자
2011. “발효 및 쿠미스,”식품 및 음료 발효 기술 핸드북,제 2 회 에딘버러. 165-172.
구글 학자
2015 년 12 월 15 일,2015 년 12 월 15 일,2015 년 12 월 15 일,2015 년 12 월 15 일,2015 년 12 월 15 일,2015 년 12 월 15 일,2015 년 12 월 15 일. 우유 발효 중 유산균 헬베티쿠스의 단백질 분해 시스템의 유전자 발현. 앤 마이크로바이올 65, 1171–1175. 10.3168.2014-8520
출판 초록|크로스레프 전체 텍스트|구글 학자
본 발명의 실시예에 따르면,본 발명의 실시예는 다음과 같다. (2011). 락토 바실러스 헬 베티 쿠스의 완전한 게놈 서열. 제.박테리올. 193, 2666–2667. 도이:10.1128/제이비.00166-11
퍼블릭 초록|크로스레프 전체 텍스트|구글 학자